Adaptation: a structure or behavior that helps an organism survive and reproduce.
Chemical: a substance that has specific characteristics based on its molecular structure.
Cue: a message that tells an organism that they should do something, or that tells them about a situation.
Gregarious: living in or spending time in groups.
Host: an organism that is carrying a parasite.
Hyperparasitoid: an insect that uses a parasitoid as a host.
Larva: the second, "worm-like" stage in the life cycle of insects that undergo complete metamorphosis (like caterpillars).
Larvae: plural of 'larva.'
Parasitize: to live in an organism as a parasite, or to make an organism a host.
Parasitoid: an insect with parasitic young that grow inside a host.
Predator: an animal that eats other animals to survive. For example, a lion is a predator... more
Prey: animal hunted for food by another animal or animals... more
Pupa: resting stage during which tissues are reorganized from larval form to adult form. The pupa is the third body form in the life cycle of insects that undergo complete metamorphosis (like caterpillars).
Signal: an act or event that sends information to someone or something.
Volatile: a chemical that changes into its gas form easily.
By Karla Moeller
Illustrated by James Baxter
Scientists communicate results through papers and often presentations. Here, anthropologist Nina Jablonski presents some of her work at a TED talk. Click for more details. Image by NASA.
One of the important things scientists do is to communicate what they learn. The common way to do this is to publish a scientific paper. These papers are usually packed with information and written for other scientists. The problem is these articles can be difficult to read.
Why "Dissect" Articles?
Scientific articles are sometimes so difficult, they seem to be written in a different language. However, lots of important information are hidden in them... information that can affect health treatments, societal views, technology developments, and more. All of that information should be able to be understood by the public. This is why we've started to break that information down, or "translate" it into a language that most people can understand.
To help get you started reading scientific papers we have dissected an article published in PLOS Biology. As you look through the paper, you’ll see that each section title has a link below it. If you click on the link, you will see the section expands with an explanation of what’s inside. We’ve also added links for each paragraph. There, we've broken down the information to be a little easier to read.
How to Read a Scientific Article
That brings us to another point: when reading a scientific paper, you don’t always want to read it from front to back. Usually, it’s helpful to skip around a bit to get the information in a certain order. Below, we’ve changed the order of sections from the original paper to improve the flow. This order can change based on why you are reading a scientific article.
Sometimes it can seem like scientific information is written in an entirely different language. Don't let it discourage you.
One of the most important things to remember as you read is to try to avoid getting discouraged. Reading scientific information can be difficult. Sometimes, you may have to re-read paragraphs two or three times before you fully understand. But if you stick with it, you will get to see a whole new world that is filled with cutting-edge information.
As a side note, you will see some numbers at the end of certain sentences. These are references. The author is telling you where they found the information in that section. To learn more, go to the References section at the end of the article.
This caterpillar is looking for food. But when it finds it, it may cause the plant to release chemical signals. Image by Sam Fraser-Smith.
At first glance, some titles can look pretty scary. But usually, they are really not that bad.
When you read, a good first rule is to look up words that you don’t know. For this title, check out the "words to know section" above for definitions of hyperparasitoid, volatile, and parasitoid.
Once you know what the words mean, you can break the title down a bit:
A plant that is being eaten by insects releases chemicals. Those chemicals can attract wasps that kill the insects. However, the wasp’s predators (another kind of wasp) can also use this chemical to find the wasps.
By Erik H. Poelman, Maaike Bruinsma, Feng Zhu, Berhane T. Weldegergis, Aline E. Boursault, Yde Jongema, Joop J. A. van Loon, Louise E. M. Vet, Jeffrey A. Harvey, and Marcel Dicke
The listed authors of an article are the people who did most of the work. Usually, the first author (here, Erik H. Poelman) was the most involved. The first author is the person you should contact if you have questions about the article. Sometimes, the last author is the senior researcher associated with the work (here, Marcel Dicke).
Some scientific articles have an Author Summary section. If they do, it’s best to read that section first (even if it is after the Abstract).
In this summary, the authors present the abstract in a way that is easier to understand.
In nature, plants often release volatiles in response to damage by herbivores (e.g., by caterpillars), and these can indirectly help defend the plants. Indeed, it is well documented that volatiles can recruit the natural enemies of herbivores, such as predators and parasitoid wasps, whose offspring feed on and develop within their caterpillar hosts. However, such induced plant odours can also be detected by other organisms. One important group of organisms, hyperparasitoids, the enemies of the parasitoids that indirectly benefit the plants, have not been included in this trophic web because so little is known about their foraging behavior. Here, using a combination of laboratory and field experiments, we demonstrate that hyperparasitoid wasps also take advantage of the odours that plants produce in response to the feeding by caterpillars. The larvae of parasitic wasps developing inside the caterpillar alter the composition of the oral secretions of their herbivorous host and thereby affect the cocktail of volatiles the plant produces. The hyperparasitoids on the lookout for their parasitoid prey can preferentially detect infected caterpillars, although not all parasitoid wasps gave away their presence through this host–plant interaction. We conclude that herbivore-induced plant volatiles can affect the interaction among parasitoids and their enemies and thereby may reduce the indirect defence accrued for the plant.
If you were bitten by someone or something, you might yell as a signal asking your neighbors for help. Image by Liam Quinn.
Oooooouuuuch!!! If someone started biting your arm, you would probably yell. A lot. This signal is how you try to get help from others and try to get your attacker to leave. Plants that get bitten can also give off signals.
When caterpillars eat a plant, the plant can give off chemicals called volatiles. Those chemicals can attract predators that eat caterpillars (like parasitoid wasps). However, the chemical can also attract other animals, including hyperparasitoid wasps. These are wasps that eat parasitoid wasps.
Scientists wanted to learn about how the plants, the caterpillars, the parasitoids, and the hyperparasitoids all interact. To do this, they ran a group of experiments both in the lab and outdoors.
When caterpillars are parasitized, it changes the signals they send to plants they eat. Click for more detail.
In this article, they share their findings with us. They show that hyperparasitoid wasps can use the plant signals to find certain caterpillars. But, how easy it is for the hyperparasitoid to find the caterpillar can depend on what the caterpillar is carrying.
If a parasitoid attacks a caterpillar, it lays its eggs inside of it, where the eggs will develop into new parasitoid wasps. Caterpillars that are carrying these wasp eggs can be easier for hyperparasitoids to find.
This means that plant chemicals can affect the relationship between these types of wasps. It also means some plants might use chemicals as a way to attract wasps, so caterpillars will stop eating the plants.
The abstract is the main summary. It usually includes the most necessary information from the whole article. In here, you will find the basics of what was already known about the subject and what was still waiting to be discovered before this project was completed.
You will also find information on the experiments the scientists did, and what they learned. With all this information, the abstract can be a little complex, so make sure to read it a few times.
Usually, it’s good to read the abstract right after the title (or right after the Author Summary). It gives you a broad sense of what is in the paper. To read it, make sure to look up words that you don’t know. It may also be good to write down notes or to draw a picture of what you read to keep it clear in your mind, as this section has a lot of information.
Plants respond to herbivory with the emission of induced plant volatiles. These volatiles may attract parasitic wasps (parasitoids) that attack the herbivores. Although in this sense the emission of volatiles has been hypothesized to be beneficial to the plant, it is still debated whether this is also the case under natural conditions because other organisms such as herbivores also respond to the emitted volatiles. One important group of organisms, the enemies of parasitoids, hyperparasitoids, has not been included in this debate because little is known about their foraging behavior. Here, we address whether hyperparasitoids use herbivore-induced plant volatiles to locate their host. We show that hyperparasitoids find their victims through herbivore-induced plant volatiles emitted in response to attack by caterpillars that in turn had been parasitized by primary parasitoids. Moreover, only one of two species of parasitoids affected herbivore-induced plant volatiles resulting in the attraction of more hyperparasitoids than volatiles from plants damaged by healthy caterpillars. This resulted in higher levels of hyperparasitism of the parasitoid that indirectly gave away its presence through its effect on plant odours induced by its caterpillar host. Here, we provide evidence for a role of compounds in the oral secretion of parasitized caterpillars that induce these changes in plant volatile emission. Our results demonstrate that the effects of herbivore-induced plant volatiles should be placed in a community-wide perspective that includes species in the fourth trophic level to improve our understanding of the ecological functions of volatile release by plants. Furthermore, these findings suggest that the impact of species in the fourth trophic level should also be considered when developing Integrated Pest Management strategies aimed at optimizing the control of insect pests using parasitoids.
If you've seen the movies Slither, Alien, or The Thing, you're pretty brave. But you're also probably familiar with the idea of one type of organism living inside of another. While this may seem creepy when it involves humans, it actually occurs regularly in the broader living world.
This ant has been parasitized by a fungus. Click for more detail.
In particular, it happens when some animals care for their young. Rather than worrying about bringing food to a baby, some mothers take baby to the food. That's exactly what parasitoid wasps do.
Parasitoids are animals that spend part of their lives inside of another type of animal, called a host. An adult parasitoid wasp lays eggs in another animal (like a caterpillar) so that her babies will have plenty of food when they hatch. The young wasps grow inside the caterpillar, using it as food.
But how do the wasps find the caterpillars? When a caterpillar eats a plant, the plant releases a specific chemical. Parasitoids can learn this signal and use it to help find the caterpillar. This creates a sort of cycle—the caterpillar eats the plant, the plant releases chemicals, and the parasitoid uses those chemicals to find the caterpillar. Because of this cycle, the plant can actually help save itself from being eaten. The chemicals can act as a signal from the plant to the parasitoid. "Help! I'm being eaten!"
However, other animals can also respond to the plant's signal. For example, other caterpillars may sense the chemicals and find the plant. After all, if a plant is already being eaten, there's a good chance it is delicious.
This hyperparasitoid wasp lays its eggs in parasitoid wasps.
But wait…we have one more level of parasite to talk about. While the parasitoids are busy looking for caterpillars to carry their young, another type of parasitoid wasp is looking for them. When a wasp lays its eggs in a parasitoid wasp, we call it a hyperparasitoid.
So how do these hyperparasitoids fit into the cycle of plant chemical, caterpillar, and parasitoid? Can they also respond to those chemicals?
In this paper, the scientists show that the chemicals given off by some plants change. Specifically, the chemicals change if the caterpillar dining on it is hosting parasitoid eggs. They also show that hyperparasitoids can respond to these changing chemicals, helping them find caterpillars hosting parasitoids.
These findings can help us understand why plants release certain chemicals. It might also help us learn to control insect pests that eat or kill plants that are especially important to us.
The intro is where you learn the background about the experiment. This not only includes information on the species being studied but it also summarizes what has been learned from other related research.
Make sure to write down notes of interest. Image by WikiHow.
In the intro, the authors usually take a look at the broader questions that are related to the study. They might explain why a particular animal may be helpful to investigate their questions, as well as what the current study will contribute to the understanding of that area of research. This section is sometimes called the "conceptual framework," as it sets up the context for the study. Most importantly, it answers the question: why is the study worthwhile?
When reading the intro, it is good to jot down notes on anything that sounds really important. Don’t forget to look up words you don't know. The authors also often briefly mention how they did the experiment and what they expected to find.
Below, we’ve added italics to what we think are the most important parts.
Plant volatiles play a profoundly important role in the structure and function of ecological communities [1]–[6]. Volatiles make a plant and its condition apparent to community members at different trophic levels [6],[7] and may, thereby, mediate interactions between organisms at higher trophic levels [8],[9]. Nowhere has this been better investigated than for interactions between insect herbivores and their natural enemies, such as primary parasitic wasps (or “parasitoids”) at the third trophic level. Many parasitoids have evolved finely tuned responses to volatiles emitted by plants that are attacked by their otherwise inconspicuous herbivorous hosts. By responding to volatiles and parasitizing the herbivores, parasitoids may reduce the amount of herbivory that plants are exposed to [1]–[3] and are, therefore, hypothesized to benefit plant fitness [10]–[12]. However, besides attracting beneficial parasitoids, the volatiles affect interactions between plants and other community members that may affect the fitness benefit of volatile release. Food webs generally include four or more trophic levels [13],[14]. Thus far, little is known about foraging behavior of the enemies of parasitoids (i.e., hyperparasitoids) that are an important group of fourth-trophic-level organisms, because hyperparasitoids have not been included in the debate on the fitness benefit of volatile release by plants [15].
Now, we've heard about plant chemicals, but why are they important? Plants use chemicals to talk, in a way. They might use them to attract animals that will pollinate them, or to warn others to stay away.
Coloration is just one way plants communicate with animals. They also release chemicals to send messages. Image by Dirk van der Made.
Depending on the chemicals released, plants can tell nearby animals different things. This may change the way that animals interact with the plant or with other animals.
For example, a caterpillar eating a plant can be hard to find. But if the plant releases a chemical when this happens, a wasp could pick up the signal and find the caterpillar.
When the wasp finds the caterpillar, it lays eggs inside, turning the caterpillar into a host. Caterpillars carrying wasp eggs are less healthy than other caterpillars and may not eat as much. So when a wasp turns a caterpillar into a host, they help stop the plant from being eaten. This helps keep the plant healthy and might be why the plant released the chemicals in the first place.
Before this study, we knew a lot about caterpillars and parasitoids. But we still needed to know how plant chemicals affect hyperparasitoid wasps.
Hyperparasitoids are parasitic wasps that attack the larvae and pupae of primary parasitoids, and they comprise a major component of the fourth trophic level in insect communities [15]. Thus far, little is known about the cues that hyperparasitoids use to find their primary parasitoid hosts [15],[16]. For secondary hyperparasitoids (i.e., hyperparasitoids that attack the fully cocooned pupae of primary parasitoids), their hosts are likely to be inconspicuous because the pupae do not feed and, therefore, do not indirectly reveal their presence through induced volatiles of the food plant. Furthermore, the time window for successful hyperparasitism of pupae is often narrow and restricted to the first few days after the pupae are formed [17]. However, plants have been shown to respond differently to feeding damage inflicted by parasitized or unparasitized herbivores [18]–[20]. As a result, plant-derived volatile cues may provide hyperparasitoids with reliable information on the presence of their host [15].
Hyperparasitoid wasps prey on the larvae (young) of parasitoid wasps. But how do they find the larvae?
Many caterpillars release chemicals into plants when they are feeding. Monarch caterpillar image by Pseudopanax.
Plants aren't the only ones that release chemicals. When caterpillars eat a plant, they release chemicals into the plant. But this chemical changes when a caterpillar is carrying wasp eggs or larvae. That, in turn, can cause plants to release a different chemical.
This means that plants release different chemicals when being eaten by a host (or parasitized) caterpillar versus a healthy one. With a different chemical comes a new message, and one that hyperparasitoids pick up. They can use this message to find parasitized caterpillars, which they use as hosts.
Many hyperparasitoid species parasitize the pupae of a range of primary parasitoid species, including both solitary species, which lay a single egg within an herbivore, and gregarious species, which lay multiple eggs within a single herbivore [21]. Single or multiple parasitoid larvae developing in a caterpillar differentially affect the physiology and feeding behavior of the herbivore, thereby inducing different plant volatile blends [19],[20]. Consequently, hyperparasitoids may be better able to locate one parasitoid host than the other, and thus variation in plant volatiles induced by parasitized herbivores may cause variation in the level of hyperparasitoid attack on different species of primary parasitoids. Here, we test whether plants can mediate interactions between third- and fourth-trophic-level organisms by providing cues on the presence of hosts for hyperparasitoids and whether hyperparasitoids respond similarly to herbivores that contain different parasitoids.
Growing up with siblings can be hard—you might argue, fight over toys or clothes, and have to share space and belongings. Growing up a single child can be tough too, though. There are benefits and drawbacks to both situations. The same is true in the animal world, so often times we see species that stick to one of those lifestyles.
Some wasps grow up in groups, like these wasps in cocoons on a hornworm caterpillar. Image by Stsmith.
Some parasitoid wasps are solitary, and almost always live alone. They are born alone, grow up alone, and often only meet with other wasps if they mate. When it is time to lay eggs, these lone wasps lay only one egg in a host. This continues the solitary cycle for their young.
Other parasitoid wasps are gregarious, spending time with a group of wasps. Gregarious adult parasitoid wasps lay a bunch of eggs in a single host. These young will grow up together and part ways once they become tiny adults.
Whether alone or in a group, parasitoid eggs hatch inside the host. Once the larvae eat enough to grow big and strong, they pupate—the process that will make them look like tiny wasps. This process takes place inside of a cocoon. So when we say cocoon, we also mean pupa.
These different sized caterpillars are healthy, but they make a good example. Host caterpillars may grow at different rates depending on how many parasites they are carrying. Image by Teles.
Though both types of these parasitoid wasps grow inside the host, they don't affect their host in the same way. At least in caterpillars, the chemicals released by a host differ if they are carrying a single wasp versus many wasps. This sends a different message to the plant. In turn, the plant sends a different message to other animals.
For hyperparasitoids, this different message might help them find either solitary or gregarious parasitoids. Scientists wanted to test out this idea.
They tested if some hyperparasitoid wasp species were more attracted to the chemicals plants release in response to caterpillars carrying one versus many wasps. How would you test to see if hyperparasitoids play favorites?
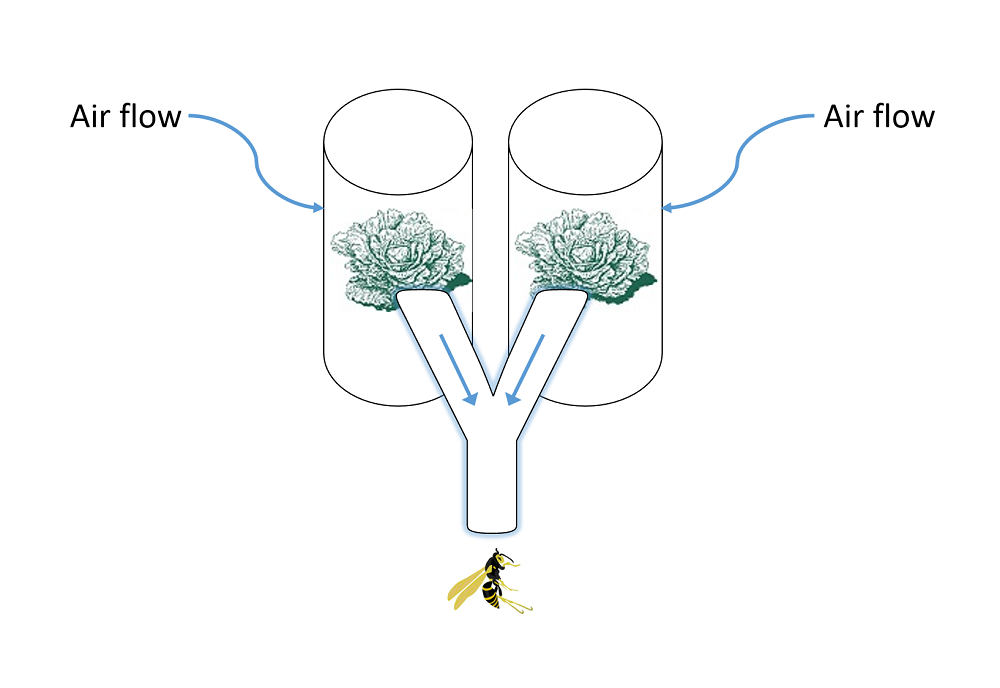
To study these questions we used the hyperparasitoid Lysibia nana (Ichneumonidae) that attacks pupae of primary parasitoids in the genus Cotesia (Hymenoptera: Braconidae). The solitary parasitoid C. rubecula (CR) and the gregarious C. glomerata (CG) are primary parasitoids that both attack caterpillars of the Small Cabbage White butterfly, Pieris rapae (PR), that feed on brassicaceous plants (Figure 1) [22]. When fully developed, the parasitoid larvae leave their host to spin a silk cocoon in which they pupate. Individual C. glomerata cocoons are approximately 40% smaller (in terms of mass) than individual C. rubecula cocoons. In terms of the per capita fitness potential of hyperparasitoid offspring, L. nana may benefit when developing in pupae of the larger C. rubecula [23]. However, in terms of cumulative maternal fitness, L. nana hyperparasitoids benefit more when finding a caterpillar parasitized by the gregarious C. glomerata . In the field, caterpillars parasitized by C. glomerata produced an average of 39 C. glomerata cocoons. Upon finding clusters of their primary parasitoid host, hyperparasitoids generally parasitize most or even all pupae in the brood, whereas in the case of finding a solitary primary parasitoid they have to disperse after each parasitization. The egg load of L. nana females closely approximates the average brood size (e.g., 20–40) of C. glomerata [17], suggesting that it co-evolved with gregarious host species such as C. glomerata . We hypothesized that L. nana uses volatile plant cues induced by C. glomerata –parasitized caterpillars to locate aggregated pupae and that they prefer those volatiles over volatile plant cues induced by the feeding of solitarily parasitized caterpillars.
The animal species used in a study are important.
The scientists used species of parasitoid wasps that prey on the larvae of the cabbage white butterfly. Image by Alvesgaspar.
Species vary, so scientists always try to pick the species that best answer their questions. In this last paragraph of the introduction, the authors discuss the types of wasps and caterpillars used in this study.
They present the species names, but we will just stick to solitary parasitoid, gregarious parasitoid, and caterpillar. Many hyperparasitoid wasp species were used, but we will treat them as one group for now. Remember that when we say those terms, we are talking about the specific species used in this study.
The scientists used one solitary and one gregarious type of parasitoid wasp. Both species feed on the caterpillars of the Small Cabbage White butterfly.
Some wasps use this long and thin ovipositor to lay eggs in caterpillars. Image by Shaw M, Jennings M, and Quicke D.
These wasps use an ovipositor—a long, sharp organ—to lay eggs inside caterpillars. The eggs turn into larvae that eat the caterpillar, then leave after they pupate.
Most hyperparasitoids only lay one egg per parasitoid larva or cocoon. But when they find a group of cocoons, they can move from one to another and lay a bunch of eggs. This means they can lay more eggs in less time and without traveling very far.
This led the scientists to guess that the hyperparasitoid wasps might prefer to lay eggs in the gregarious wasp cocoons. If they do prefer the gregarious wasps, then they will likely use the signals from plants to find those larvae or cocoons.
Before going to the next full section, it’s good to briefly look over some of the figures. This helps you get a better picture in your head of the information included in the paper and of what the scientists did. When you first review the images, it's good to focus on just one or two that help set up the ideas behind the experiment.
Click to enlarge.
Figure 1. Experimental study system of the four-trophic-level community on Brassica oleracea plants. The gregarious primary parasitoid Cotesia glomerata (CG) and the solitary C. rubecula (CR) attack caterpillars of Pieris (PR) butterflies, which are in turn attacked by several hyperparasitoids: Acrolyta nens (1), Lysibia nana (2), Pteromalus semotus (3),Mesochorus gemellus (4), and Baryscapus galactopus (5). Hyperparasitoids at the fourth trophic level find their primary parasitoid host at the third trophic level via information derived from the plant at the first trophic level. Larvae of primary parasitoids that develop in their herbivorous host at the second trophic level inflict changes in their herbivore host, and the combination of herbivore and parasitoid (parasitized herbivores) inflict changes in plant volatile emission (I). These changes in plant volatile emission are used by hyperparasitoids as a cue of host presence (II). Photograph credit: Tibor Bukovinszky. doi:10.1371/journal.pbio.1001435.g001
Figure 1 is a map of the scientists' story. This figure shows us the different species involved in this story and how they relate to one another.
We see the plant at the bottom of the picture. Then, with each step up, we are looking at animals that eat the organism below it.
Above the plant is the caterpillar, which eats the plant. Above the caterpillar we see two parasitoid wasps. The one on the right is the solitary wasp and the one on the left is the gregarious (or grouping) wasp.
Here, we see a wasp using its ovipositor to lay an egg in a caterpillar. Neither of these species were used in this study. Image by USDA Agricultural Research Service.
Both of these wasps lay their eggs in the caterpillar (you can see the eggs and larvae in the picture).
Above the parasitoid wasps are five kinds of hyperparasitoid wasps. These wasps all lay their eggs in both kinds of parasitoid wasp larvae or cocoons.
These different layers of predators are called trophic levels. Trophic just means related to food, so you can think of them as the level at which an animal eats in its food chain. The plant is the first food, or trophic, level. Each step up adds another trophic level, so that the top of the picture is the fourth trophic level.
This figure also talks about some of the results that the scientists found. When the parasitoids lay eggs in the caterpillars, it changes the way the caterpillars affect the plants. These changes cause the plant to release a different chemical, which helps attract the hyperparasitoid. We will review this finding again in other figures and results.
Click to enlarge.
Figure 4. Preference of hyperparasitoids for herbivore-induced plant volatiles. Preference of the hyperparasitoid Lysibia nana for herbivore-induced plant volatiles was tested by using a full factorial design of two-choice olfactometer tests including pair-wise comparisons of the treatments: undamaged plants (white bars), Pieris rapae damaged plants (light grey), plants damaged by Pieris rapae caterpillars parasitized by Cotesia glomerata (dark grey bars), or plants damaged by Pieris rapae caterpillars parasitized by C. rubecula (black bars). The two lowest pairs of bars show the preference of L. nana for plants treated with caterpillar regurgitant. The first pair shows hyperparasitoid preference when plants are artificially damaged and regurgitant of unparasitized (light grey) or parasitized (dark grey) caterpillars was applied. The second and lowest pair shows that hyperparasitoids do not respond to the application of regurgitant without artificially damaging the plant. Numbers between brackets indicate the number of wasps that made a choice within 10 min from the start of the experiment versus the total number of wasps tested. * p<0.05, ** p<0.001. Photograph credit: Tibor Bukovinszky. doi:10.1371/journal.pbio.1001435.g004
Caterpillars are a favorite food for some wasps. Image by Benny Mazur.
We all have favorite foods. It turns out that wasps do as well. Figure 4 shows us what conditions and chemicals most attract hyperparasitoids. This is a choice experiment.
Wasps were presented with two choices. Each choice is represented by a color on a paired bar. Whichever way the bar is shifted shows which choice the wasps preferred.
The scientists tested whether one species of hyperparasitoid wasp was more attracted to plants from any of four different groups. The groups were as follows.
Uneaten plants.
Plants eaten by healthy caterpillars.
Plants eaten by parasitized caterpillars carrying one larva from the solitary wasp.
Plants eaten by parasitized caterpillars carrying many larvae from the gregarious wasp.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
They also checked to see if wasps liked damaged plants that were painted with caterpillar throw up. When the paired bars are split pretty evenly by the middle line, it means the hyperparasitoid wasp didn't prefer either group. When the paired bars are more to one side and have one or two stars (asterisks), the hyperparasitoid wasp preferred one of the two paired groups.
The top three pairs of bars compare the wasps' attraction to different plant treatments. The healthy, uneaten leaves are shown by the white sides of the bars. Leaves that were damaged by three different caterpillar groups are shown by the grey and black bars. The light grey bar on top shows leaves damaged by healthy caterpillars. The dark grey bar in the middle shows leaves damaged by caterpillars hosting many gregarious larvae. The black shows leaves damaged by caterpillars hosting one solitary larva.
Wasps liked plants that had been damaged by caterpillars more than they liked healthy plants. Image by MukherjeeApratim.
All three pairs of these bars are shifted very far to the right. This means the hyperparasitoid wasps were attracted to plants damaged by all three caterpillar groups.
The middle three pairs of bars compare the damaged plant treatments to each other. The top and middle bars show plants damaged by healthy caterpillars (light grey bars) versus parasitized caterpillars. Caterpillars with gregarious larvae are shown on the top bar (dark grey). Caterpillars with solitary larva are shown on the middle bar (black). The bottom paired bar of this set compares the two types of parasitized caterpillars. Those carrying solitary larva are shown in black. The bars are all shifted toward the dark grey, gregarious treatments. This means the hyperparasitoid wasps favored plants damaged by caterpillars carrying many larvae.
At this point, the scientists knew the wasps had a preference, but what was it they were attracted to?
Scientists learned that wasps most preferred plants damaged by caterpillars carrying gregarious larvae. Image by Stsmith.
To figure this out, they looked at how caterpillar throw up (called regurgitant) affected wasp attraction. The pair of bars that is second from the bottom compares plants damaged by people and treated with caterpillar throw up. The light grey on the left shows healthy caterpillar throw up. The dark grey on the right shows throw up from caterpillars hosting many larvae. Even though the caterpillar hadn't directly touched the plant, the wasps still showed preference for the parasitized caterpillars.
The scientists still had to test which chemicals were attracting the wasps: the plant chemicals? Or the caterpillar chemicals? The lowest pair of bars compares the chemicals in caterpillar throw up to the chemicals the plants release. But in this case, the plants were not damaged. When plants were undamaged, the wasps had no preference for either plant, showing the throw up by itself was not what attracts the wasps. Instead, wasps are attracted to the chemicals the plant gives off in response to the caterpillar throw up.
In school, you probably learned to write science reports in a very specific way. First comes the introduction (where you discuss hypotheses), then the methods (where you discuss what you did). Only after that do you get to the results (where you discuss what you found).
The results can be considered the heart of the paper. It is the main paper part that keeps information flowing to other scientists. Image by GustavoHCL.
However, in many scientific articles, the methods are saved for the end. This is because unless you are interested in a specific method, that section won't tell you a whole lot about the subject.
So, once we get through the introduction, we are suddenly at the results section. This takes us right to the heart of what the scientists found.
Usually, if there are a lot of results, they will be split up into subsections to help you find the results that interest you. Here, we will go through all of the results. We will also present the figures close to where the authors discuss them.
In line with differences in the size of their hosts, we found that L. nana wasps were about 35% smaller when emerging from C. glomerata than C. rubecula cocoons (Table 1; Figure 2).
To start off the results, the scientists note that the choice of parasitoid wasp host has a big effect on a hyperparasitoid wasp's young. Solitary larvae are larger, so wasps laid in solitary larvae can grow to be larger as well.
Even though gregarious cocoons were smaller, wasps preferred them to solitary cocoons. Image by Hans Hillewaert
When most animals are young, being larger is better, as it makes it a bit easier to survive. However, as we saw in Figure 4, hyperparasitoid wasps were more attracted to the gregarious wasps. We will talk about this unexpected finding more in the discussion.
We won't go over Figure 1 again because it didn't present any results. Instead we will go next to Figure 2 and Table 1. They are associated with the findings discussed above.
Click to enlarge.
Figure 2. Performance of Lysibia nana on pupae of two parasitoid species.Lysibia nana dry mass plotted against the mass of the Cotesia cocoon before L. nana had parasitized the cocoon. Orange symbols represent wasps emerging from C. glomerata cocoons, and black symbols those emerging from C. rubecula cocoons. Females are represented by dots, and males by triangles. Photograph credit: Tibor Bukovinszky. doi:10.1371/journal.pbio.1001435.g002
Size and space are closely related.
Think of a chicken egg, for example. If you have a large egg and a much smaller egg that are both about to hatch, which do you think will have a larger chick?
Larger young will often come out of larger eggs. This is because the larger egg usually has more resources the developing young can use to grow. Image by Batholith.
The larger egg, right? It would be impossible for a larger chick to fit into the much smaller egg. Egg or cocoon size controls the size of young in other animals as well.
This graph shows the size of wasps coming out of, or emerging from, a range of cocoon sizes.
On the vertical (left) axis, we see the mass of young hyperparasitoid wasps when they come out of the cocoons of parasitoid wasps. On the horizontal (bottom) axis, we see the mass of the cocoons.
Orange marks show hyperparasitoid wasps that came out of smaller, gregarious wasp cocoons. Black marks show hyperparasitoid wasps that came out of larger solitary wasp cocoons.
Wasps that grew in smaller cocoons ended up smaller than wasps that grew in larger cocoons.
Click to enlarge.
Table 1. Fresh weight and clutch size of cocoons of two Cotesia species collected from a laboratory rearing or from the field, and the corresponding fitness-related traits in the hyperparasitoid Lysibia nana.
doi:10.1371/journal.pbio.1001435.t001
Table 1 shows us the average of the masses of cocoons and wasps shown above in Figure 2.
When testing the response to volatiles directly derived from cocoons in a Y-tube olfactometer,L. nana wasps were not attracted to odours associated with the pupae, such as those derived from silk with which the wasps have spun a cocoon to house the pupa. Only 5% of the wasps tested walked up the olfactometer arm within 10 min. Therefore, we studied L. nana responses to cocoons when offered at a closer range, similar to conditions that hyperparasitoids may experience after landing on a plant. L. nana females were offered a choice between a brood of the gregarious parasitoid C. glomerata (CG) and a cocoon of the solitary wasp C. rubecula (CR) in a glass Petri dish (18.5 cm diameter, 4.6 cm height). When the hyperparasitoid wasps were released, we did not observe any directional movement towards the cocoons and the hyperparasitoid wasps often passed cocoons within a centimeter distance. Despite their activity and limited search area, within 10 min, 45 of the 70 tested wasps encountered one of the cocoons and were arrested. A cocoon clutch of C. glomerata was more commonly found than a solitary cocoon of C. rubecula (binomial test, n = 45, p<0.001) (Figure 3). When we offered cocoons of C. rubecula in a group, to match the cocoon mass of a brood of C. glomerata cocoons, we found that L. nana females more often first encountered the group of C. rubecula cocoons than the clutch of C. glomerata cocoons (n = 47, p = 0.003) (Figure 3). This may be due to the larger surface covered by the group of C. rubecula cocoons than the surface covered by the brood of C. glomerata cocoons.
The scientists tested if the hyperparasitoid wasps preferred one of the parasitoid species. When allowed to choose based on scent alone, the wasps had no preference. Yet, when they found cocoons, they usually found those of the gregarious wasps.
Eyesight may be more important than scent for some wasps. Image by the USGS Native Bee Inventory and Monitoring Laboratory.
The scientists tested further to see if maybe the wasps were using scent at close range. They also thought the wasps might be choosing based on the larger size of the grouped cocoons.
When several solitary cocoons were grouped together, the wasps found them quicker. This suggests that the wasps were using size rather than scent to choose. They chose whichever group of cocoons was easier to spot.
Click to enlarge.
Figure 3. Lysibia nana responses in choice tests with primary parasitoid cocoons.
Lysibia nana preference (top bar) for gregarious broods of Cotesia glomerata (grey) or solitary cocoons of C. rubecula (white) in a Petri dish bioassay. Lysibia nana preference (lower bar) for gregarious broods of Cotesia glomerata (grey) or the same number of cocoons of C. rubecula (white). Numbers between brackets indicate the fraction of wasps that responded to cocoons within 10 min from the start of the experiment. *p<0.05, ** p<0.001. Photograph credit: Tibor Bukovinszky. doi:10.1371/journal.pbio.1001435.g003
Does size matter? The scientists decided to test directly whether hyperparasitoids preferred one type of parasitoid all the time, or if it was based on size of larvae or cocoons.
Wasps were released into petri dishes that held a gregarious group of cocoons and either a solitary cocoon or a group of solitary cocoons. Petri dish by Lilly_M.
Figure 3 shows paired bars, with the bars shifted to one side. This shows a preference of the hyperparasitoid wasps for the cocoons on that side.
On the left, we see the yellow group of small cocoons of the gregarious parasitoid wasps. On the right, we see the cocoons of the solitary parasitoid wasp. The lower right picture is of solitary cocoons that the scientists placed together in a group.
Groups of these different cocoons were placed in small Petri dishes (like you may see people use to grow bacteria). Wasps were released into the dish. The cocoon type that was found first by the wasp was recorded. The bars show us the percentage of time the wasps chose (or found) one cocoon type.
Gregarious parasitoid cocoons often take up more space than solitary eggs. Image by GothMoths.
As you can see, on the top, when the gregarious cocoons took up more space, the hyperparasitoid wasps found them more quickly. However, when the solitary cocoons were grouped together to cover more space, the wasps found these more quickly.
These findings suggest that the wasps had no preference for the chemicals (scent) given off by either of the cocoon types. Instead, they seem to have found whichever cocoon type was taking up more space.
Response of Hyperparasitoids to Herbivore-Induced Plant Volatiles
Hyperparasitoids did respond to herbivore-induced plant volatiles. L. nana females preferred volatiles from plants damaged by either caterpillars parasitized by primary parasitoids (both C. glomerata [PR-CG] and C. rubecula [PR-CR]) or unparasitized caterpillars (PR) over volatiles from undamaged plants in a Y-tube olfactometer (Figure 4; binomial tests, p<0.001). The hyperparasitoids did not discriminate between volatile blends from plants damaged by unparasitized caterpillars and plants damaged by caterpillars parasitized by the solitary parasitoid C. rubecula (Figure 4; binomial test, p = 0.480). However, plant volatiles induced by C. glomerata–parasitized caterpillars were more attractive to L. nana than volatiles from plants damaged by unparasitized caterpillars or those parasitized by the solitary parasitoid C. rubecula (binomial tests, p = 0.021 and p = 0.007, respectively) (Figure 4). Unparasitized and C. glomerata–parasitized caterpillars inflicted more damage to plants than caterpillars parasitized by a solitary C. rubecula parasitoid—that is, respectively, 2,485±1,183 (mean ± SD), 1,855±810, and 705±313 mm2 of leaf tissue consumed per caterpillar in 24 h (Kruskal-Wallis Test, p<0.001). Because parasitoid species differentially affect caterpillar feeding rate by regulating the growth of their host [19] and the rate of feeding damage per se may mediate the attraction of L. nana, we controlled for the amount of damage in a subsequent test. We damaged plants with a pattern wheel and applied 25 µl of oral secretion from either unparasitized or parasitized caterpillars to the damaged sites. Parasitoid species are known to alter the composition of the oral secretions of their host and thereby strongly affect the response of the plant to a parasitized caterpillar [20]. Compounds in the oral secretions of Pieris caterpillars play a key role in inducing volatile release by their food plant [24]. L. nana preferred the volatiles from plants that were treated with oral secretions obtained from C. glomerata–parasitized caterpillars over volatiles from plants treated with oral secretions from unparasitized caterpillars (Figure 4). Oral secretions of parasitized caterpillars alone (i.e., without application to wounded sites) did not attract the hyperparasitoids (Figure 4).
Hyperparasitoid wasps prefer certain parasitoid wasps. But what helps them tell the difference between wasps? Chemicals from the caterpillars that host the wasps? Or from the plants?
Hyperparasitoid wasps preferred plants that had been damaged. Image by Primejyothi.
The wasps weren't reacting to chemicals from the caterpillars. Instead, they were reacting to chemicals from the plants. They especially liked plants that had been damaged.
But the type of damage was key. The wasps didn't like plants damaged by healthy caterpillars. They also didn't care much for plants damaged by caterpillars hosting solitary larvae. However, the wasps did like plants damaged by caterpillars hosting gregarious larvae.
Could this be due to the damage the caterpillars cause? Depending on what type of parasitoid a caterpillar carries, they may eat more or less. So, in order to figure out what was really going on, the scientists controlled the plant damage.
Scientists needed to control the amount of damage caterpillars were doing. They compared plant damage that was controlled to that inflicted by caterpillars and to undamaged plants.
The scientists damaged the plants. Then they put oral secretions (basically caterpillar throw up) on the plant's wounds. This made it so the caterpillars couldn't affect the findings.
Wasps preferred plants that had been treated with throw up from caterpillars carrying gregarious larvae. This showed that the wasp's preference wasn't due to caterpillar behavior. Instead, it was due to the chemicals the plants released in response to the caterpillar throw up.
Now, let's review Figure 4 again, because it explains these findings.
Click to enlarge.
Figure 4. Preference of hyperparasitoids for herbivore-induced plant volatiles.
Preference of the hyperparasitoid Lysibia nana for herbivore-induced plant volatiles was tested by using a full factorial design of two-choice olfactometer tests including pair-wise comparisons of the treatments: undamaged plants (white bars), Pieris rapae damaged plants (light grey), plants damaged by Pieris rapae caterpillars parasitized by Cotesia glomerata (dark grey bars), or plants damaged by Pieris rapae caterpillars parasitized byC. rubecula (black bars). The two lowest pairs of bars show the preference of L. nana for plants treated with caterpillar regurgitant. The first pair shows hyperparasitoid preference when plants are artificially damaged and regurgitant of unparasitized (light grey) or parasitized (dark grey) caterpillars was applied. The second and lowest pair shows that hyperparasitoids do not respond to the application of regurgitant without artificially damaging the plant. Numbers between brackets indicate the number of wasps that made a choice within 10 min from the start of the experiment versus the total number of wasps tested. * p<0.05, ** p<0.001. Photograph credit: Tibor Bukovinszky. doi:10.1371/journal.pbio.1001435.g004
Caterpillars are a favorite food for some wasps. Image by Benny Mazur.
We all have favorite foods. It turns out that wasps do as well. Figure 4 shows us what conditions and chemicals most attract hyperparasitoids.
This is a choice experiment. Wasps were presented with two choices. Each choice is represented by a color on a paired bar. Whichever way the bar is shifted shows which choice the wasps preferred.
The scientists tested whether one species of hyperparasitoid wasp was more attracted to plants from any of four different groups. The groups were as follows.
Uneaten plants.
Plants eaten by healthy caterpillars.
Plants eaten by parasitized caterpillars carrying one larva from the solitary wasp.
Plants eaten by parasitized caterpillars carrying many larvae from the gregarious wasp.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
They also checked to see if wasps liked damaged plants that were painted with caterpillar throw up.
When the paired bars are split pretty evenly by the middle line, it means the hyperparasitoid wasp didn't prefer either group. When the paired bars are more to one side and have one or two stars (asterisks), the hyperparasitoid wasp preferred one of the two paired groups.
The top three pairs of bars compare the wasps' attraction to different plant treatments. The healthy, uneaten leaves are shown by the white sides of the bars. Leaves that were damaged by three different caterpillar groups are shown by the grey and black bars. The light grey bar on top shows leaves damaged by healthy caterpillars. The dark grey bar in the middle shows leaves damaged by caterpillars hosting many gregarious larvae. The black shows leaves damaged by caterpillars hosting one solitary larva.
Wasps liked plants that had been damaged by caterpillars more than they liked healthy plants. Image by MukherjeeApratim.
All three pairs of these bars are shifted very far to the right. This means the hyperparasitoid wasps were attracted to plants damaged by all three caterpillar groups.
The middle three pairs of bars compare the damaged plant treatments to each other. The top and middle bars show plants damaged by healthy caterpillars (light grey bars) versus parasitized caterpillars. Caterpillars with gregarious larvae are shown on the top bar (dark grey).
Caterpillars with solitary larva are shown on the middle bar (black). The bottom paired bar of this set compares the two types of parasitized caterpillars. Those carrying solitary larva are shown in black. The bars are all shifted toward the dark grey, gregarious treatments. This means the hyperparasitoid wasps favored plants damaged by caterpillars carrying many larvae.
Scientists learned that wasps most preferred plants damaged by caterpillars carrying gregarious larvae. Image by Stsmith.
At this point, the scientists knew the wasps had a preference, but what was it they were attracted to? To figure this out, they looked at how caterpillar throw up (called regurgitant) affected wasp attraction.
The pair of bars that is second from the bottom compares plants damaged by people and treated with caterpillar throw up. The light grey on the left shows healthy caterpillar throw up. The dark grey on the right shows throw up from caterpillars hosting many larvae. Even though the caterpillar hadn't directly touched the plant, the wasps still showed preference for the parasitized caterpillars.
The scientists still had to test which chemicals were attracting the wasps: the plant chemicals? Or the caterpillar chemicals? The lowest pair of bars compares the chemicals in caterpillar throw up to the chemicals the plants release. But in this case, the plants were not damaged. When plants were undamaged, the wasps had no preference for either plant, showing the throw up by itself was not what attracts the wasps. Instead, wasps are attracted to the chemicals the plant gives off in response to the caterpillar throw up.
Volatiles
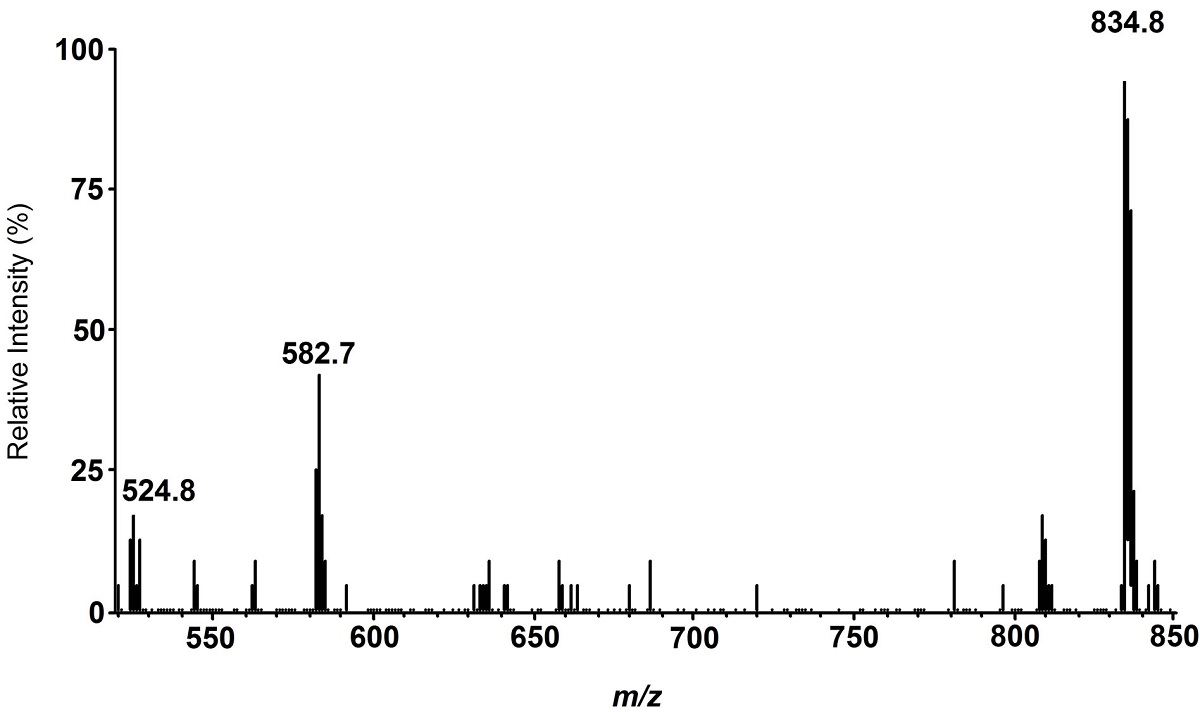
Analysis of the volatile blends of plants induced by C. glomerata–parasitized, C. rubecula–parasitized, or unparasitized caterpillars revealed that these three herbivore treatments induce volatile blends that differ from undamaged control plants. A total of 33 compounds that were present in all samples of at least one of the four plant treatments were tentatively identified and included in further analysis (Table 2). In the PLS-DA, undamaged control plants grouped separately from the three treatments with caterpillar feeding (Figure 5). Amongst the caterpillar-damage treatments, plants damaged by feeding of unparasitized caterpillars and caterpillars parasitized by C. rubecula overlapped largely in their volatile headspace as shown by PLS-DA. Plants damaged by C. glomerata–parasitized caterpillars were only 40% similar in their volatile headspace to plant headspaces induced by the two other caterpillar treatments and were most distinctly different from undamaged control plants. Nine compounds most strongly contributed to the differences among treatments are indicated by VIP scores higher than 1. These compounds included terpenoids, a ketone, a nitrile, and two unknown compounds (Table 2). The concentrations of two compounds differed significantly among the caterpillar treatments. Plants damaged by C. glomerata–parasitized caterpillars produced higher concentrations of (E)-4,8-dimethylnona-1,3,7-triene [(E)-DMNT], a known attractant for parasitoids [25], and of an unknown compound compared to plants damaged by C. rubecula–parasitized or unparasitized caterpillars. The similarity of the volatile blends from plants damaged by unparasitized and C. rubecula–parasitized P. rapae matches the observation that hyperparasitoids did not discriminate the two treatments in choice assays. The hyperparasitoids did prefer plants damaged by C. glomerata–parasitized caterpillars over other damage treatments, which is supported by the difference in the composition of the volatile blends emitted by the plants submitted to these treatments.
The hyperparasitoid wasps use plant signals to find parasitoids. But what is in the plant signals? When being eaten by caterpillars, the plants released 33 different chemicals. Using a special test, the scientists found which chemicals were most different. Nine chemicals were most important to the changing signals released by the plants.
The same plant can release different chemical signals, changing the message sent to other organisms. Image by Rajeevvadakkedath.
The undamaged plants released very different chemicals from all of the damaged plants. Plants damaged by healthy caterpillars and those hosting solitary wasps released a lot of the same chemicals. This may explain why the hyperparasitoids didn't prefer one of those two signals.
But plants damaged by caterpillars hosting gregarious wasps were different. They released a different chemical mix. This is what attracted the wasps.
Another way these plants changed their signal was by altering the amount of chemicals they released. Caterpillars with gregarious parasitoids caused plants to release lots of chemicals.
Click to enlarge.
Figure 5. PLS-DA plot based on comparisons among volatile blends of Brassica oleracea plants under herbivory by parasitized or unparasitized caterpillars.
Plants were either undamaged (red, UD), damaged with two unparasitized Pieris rapae caterpillars (blue, PR), or P. rapae caterpillars parasitized by Cotesia rubecula (orange, PR-CR) or C. glomerata (green, PR-CG). Photograph credit: Tibor Bukovinszky. doi:10.1371/journal.pbio.1001435.g005
At first, this graph probably looks like a bunch of messy shapes and colors. But it shows the different chemical mixes released by plants in the study. Most importantly, it shows how these chemical mixes overlap and differ.
We often think of liquid chemicals in laboratories, but in this study, scientists analyzed chemicals in gas form. Image by RIA Novosti.
The red outline shows the range of chemicals released by undamaged plants. You can see that this group of chemicals has the least amount of overlap with the others.
The green is the most different from the undamaged plants. You can see there is no overlap. The lines of the two shapes don't even touch. Green shows the chemicals from plants that were damaged by caterpillars hosting gregarious wasps. The different chemicals allowed hyperparasitoid wasps to find these caterpillars.
The blue area shows the chemicals released by plants that were damaged by healthy caterpillars. It overlaps almost entirely with parts of the yellow area. The yellow area shows chemicals released by plants damaged by caterpillars hosting solitary wasps. The overlap between these two chemical mixes would likely make it difficult for hyperparasitoids to tell the difference between them.
Click to enlarge.
Table 2. Volatile compounds detected in the headspace of Brassica oleracea, uninfested (control), or infested with two caterpillars of Pieris rapae that were either unparasitized or parasitized with Cotesia rubecula or C. glomerata.
doi:10.1371/journal.pbio.1001435.t002
This table shows the list of chemicals the plants released.
Scientists collected gaseous chemicals from the space around plants from different treatments. Image by Hannah 50.
At the top of the table, you see the list of treatments the scientists gave the plant. First they list the undamaged plants, also called the control. Next are the plants damaged by healthy caterpillars. The two other treatments were plants damaged by caterpillars parasitized by either one of the two parasitoids.
Chemicals were released into what scientists call the plant's headspace, or the air around the plant. Across all the plants, in this air, 33 different chemicals were found. Chemicals that were most different between treatments were considered more important. This is shown with the VIP score (on the right). The VIP scores are higher and in bold for the chemicals that were most different between the treatments.
Hyperparasitoid Responses in the Field
In the field, we confirmed that plant volatiles play an important role in the location of parasitoid pupae by hyperparasitoids. In an experimental field, B. oleracea plants were subjected to four induction treatments: no damage (UD), feeding by healthy P. rapae caterpillars (PR), or feeding by P. rapae caterpillars parasitized by either C. rubecula (PR-CR) or C. glomerata (PR-CG). After the caterpillars had fed on the plants for 10 d, which was approximately the total development period of the Cotesia larvae, the caterpillars were removed. On half of the plants per treatment, we then attached C. glomerata cocoons and on the other half C. rubecula cocoons. The cocoons were exposed to the natural population of hyperparasitoids and recollected to assess the number of cocoons that was hyperparasitized. C. glomerata pupae that were attached to plants damaged by C. glomerata–parasitized caterpillars were more frequently hyperparasitized than pupae attached to plants damaged by unparasitized or C. rubecula–parasitized caterpillars (Figure 6, Table 3). However, when C. rubecula cocoons were used to assess hyperparasitism rates, we found no induction treatment effect.
Scientists did a lab experiment to check if what they saw in the lab happened in nature. Image by Quahadi Añtó.
If possible, experiments are done both in a science lab and in the field (in the wild). This helps to make sure that what we see in lab actually happens in nature.
These scientists did a field experiment to check some of what they learned in the lab. They wanted to make sure that hyperparasitoids use plant chemicals to find parasitoid hosts in the field.
They used the same four plant treatments:
Undamaged.
Damaged by healthy caterpillars.
Damaged by caterpillars hosting solitary parasitoids.
Damaged by caterpillars hosting gregarious parasitoids.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
The caterpillars were allowed to feed on the plants for ten days and then they were removed.
Caterpillar cocoons with parasitoids were then put on plants from each treatment group. This made it so that all different plant chemicals would be matched with both of the two parasitoids. Half of the plants in each group had solitary parasitoids. The other half of the plants had gregarious parasitoids.
Scientists removed the caterpillars from the damaged plants and attached parasitoid cocoons before uncovering them. This way they knew that local hyperparasitoid preference was not affected directly by the caterpillars. Image by Rasbak.
The cocoons were then left in the open for nearby hyperparasitoids to find them. This way, the scientists could see how important plant chemicals were to hyperparasitoid preference in the field.
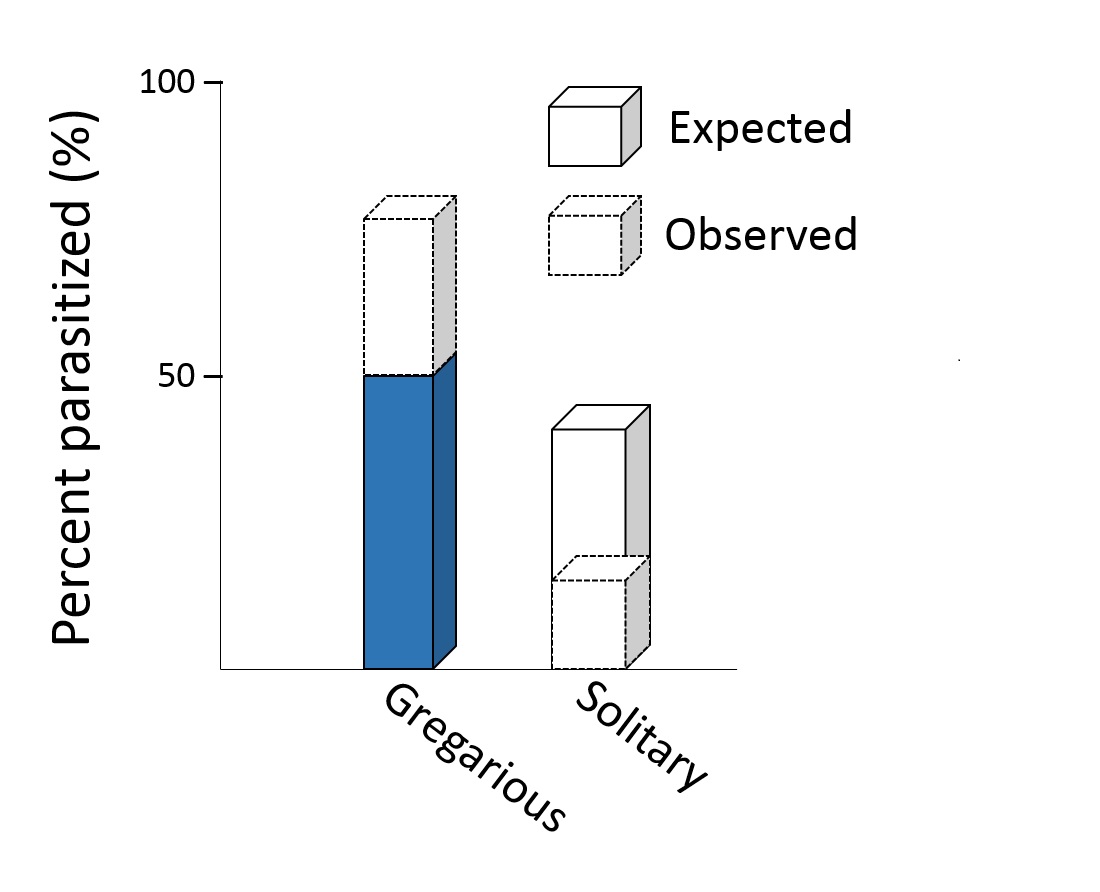
Plant group had an effect on wasp preference for gregarious parasitoids. The wasps laid the most eggs in gregarious parasitoids that were on plants damaged by hosts of the same parasitoid. Next in order of preference were gregarious parasitoids on any damaged plants. They laid the least eggs in hosts on undamaged plants.
With solitary parasitoids, plant group did not change wasp preference. This means that the plant chemicals may be more important to wasps looking for gregarious hosts.
Click to enlarge.
Figure 6. Herbivore-induced plant volatiles mediate hyperparasitism in the field.
Percentage of Cotesia glomerata (CG, left) and C. rubecula (CR, right) cocoon clutches hyperparasitized on plants that had been induced with herbivory by unparasitized or parasitized caterpillars of P. rapae. Pieris rapae (PR), P. rapae parasitized by C. glomerata (PR-CG), P. rapae parasitized by C. rubecula (PR-CR), and undamaged (UD). Letters indicate significant differences between treatment groups (GLM, p<0.05). Photograph credit: Tibor Bukovinszky.
doi:10.1371/journal.pbio.1001435.g006
This graph shows the results from the field experiment.
On the bottom (x-axis) of the graph, we see the same four plant treatments:
Undamaged.
Damaged by healthy caterpillars.
Damaged by caterpillars hosting solitary parasitoids.
Damaged by caterpillars hosting gregarious parasitoids.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
The treatments are split into two groups. Within each group, the height of the bar shows the percentage of cocoons in which hyperparasitoids laid eggs.
The bars on the left show gregarious parasitoid cocoons attached to plants from each treatment. The bars on the right show the solitary parasitoid cocoons attached to plants from each treatment. The two groups were not compared directly.
On the left side, we see that the tallest bar (labeled "c") represents gregarious cocoons that were placed on plants that had been damaged by hosts of the same parasitoid. These cocoons were favored by the hyperparasitoid wasps.
Wasps most preferred gregarious cocoons that were on plants damaged by caterpillars carrying gregarious parasitoids. Image by Hans Hillewaert.
The letters above the bars indicate how different the groups were from each other. Bars that have the same letter (b, for example) show groups that had similar attack rates from hyperparasitoids. If the letters differ, this means the groups attracted different numbers of hyperparasitoids.
Next in preference for the gregarious group were cocoons on any damaged plants. Least favored were gregarious cocoons on undamaged plants. In the solitary parasitoid group, plant treatment did not change wasp preference. We can tell this because there are no letters to show differences between the groups.
We will talk about these results in more depth in the discussion section.
Click to enlarge.
Table 3. The effect of plant induction treatment on the fraction of primary parasitoid cocoons per plant that contained any hyperparasitoid in the field.
doi:10.1371/journal.pbio.1001435.t003
In order to use the data collected in an experiment, we must use statistics. This table shows the statistical results of the field experiment. Let's talk about the terms the scientists used before we talk about statistics.
The two main factors are listed here as caterpillar induction and type of cocoon. Caterpillar induction just refers to the same plant treatment groups we talked about above:
Undamaged.
Damaged by healthy caterpillars.
Damaged by caterpillars hosting solitary parasitoids.
Damaged by caterpillars hosting gregarious parasitoids.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
Type of cocoons describe which parasitoid was added to the plants after they were damaged by the caterpillars.
When we do statistical tests in biology, we are testing to see whether or not an outcome can be explained by chance. If it isn't explained by chance, it is likely explained by one of the factors that we are testing. The P value shows us how probable it is that we would get our results if there was no connection between a factor and our findings. This number is always between zero and one.
If there is no relationship, we would expect a higher P value (close to one). This would mean that our findings are likely due to chance. The lower the P value (closer to zero), the more rare our results. So lower P values mean the factor we are looking at affected our findings.
When scientists run statistics, they are comparing data they observed from an experimental group to either predicted data or data from a control group. Curves that overlap a lot are considered to be the same, so the treatment had no real effect.
As a general rule, scientists say that a factor has an effect if the P value is less than 0.05. This means there is a 5% or less probability that we would get the observed result if it was due to chance.
Here we see that caterpillar induction (plant treatment) and type of cocoon both have P values below 0.05. This means that both of these factors are important to hyperparasitoids when they are deciding where to lay their eggs.
The second factor that was measured, replicate, deals with variation. Variation tells us how much an individual or a group differs from one measurement to the next. In the table, we see that replicate had a very low P value. This means that the cocoon-plant treatment groups varied quite a bit even when the scientists tried to replicate them.
Both cocoon type and plant treatment were important to hyperparasitoid preference. Image by Doug Beckers.
In the lower part of the graph, you see that the interaction between factors has also been measured. This tells us how much the factors depend on each other. So, for example, induction and cocoon type have a significant (this is what we say when the P value is less than 0.05) interaction.
This means that having both of these factors present is important to get the results the scientists observed. Basically, this shows us that the effect of the cocoon type on the hyperparasitoids depends on what plant treatment was given (and vice versa).
The preference of L. nana for volatiles derived from plants damaged by C. glomerata–parasitized caterpillars has profound consequences for the primary parasitoid C. glomerata in the field. During the growing season of cabbage plants in 3 consecutive years in the vicinity of Wageningen, the Netherlands, we collected 1,256 cocoon clusters of the gregarious primary parasitoid C. glomerata and 1,668 cocoons of the solitary primary parasitoid C. rubecula and assessed the rate of natural hyperparasitoid attack. Clusters of C. glomerata cocoons more often contained at least a single hyperparasitoid than did solitary cocoons of C. rubecula(Generalized Linear Model, deviance = 496.62, p<0.001; Table 4). From 17.4% of the C. glomerata clusters, more than one (and occasionally even four) hyperparasitoid species emerged. Hyperparasitoid communities associated with the gregarious primary parasitoid also consisted of more species than were found on the solitary parasitoid (Figure 7; Table 5). Within clusters of C. glomerata cocoons that were attacked by hyperparasitoids, 65%–81% of the pupae in the cluster yielded hyperparasitoid wasps. The combined attack rate of clusters and the fraction of pupae hyperparasitized in a cluster resulted in a total hyperparasitism rate of individual C. glomerata pupae of 20%–55% over the 3 y (Figure 7). By contrast, only 5%–15% of individual C. rubecula cocoons were hyperparasitized over the 3 y of our field experiments. Gregarious C. glomerata are, therefore, not only more easily found by hyperparasitoids, but once found, the hyperparasitoid wasps are able to parasitize large numbers of parasitoid pupae within broods, revealing that they exhibit an aggregative response to this clustered resource.
The scientists also did a study to see the attack rates of hyperparasitoids in the field. For this, they had to collect a lot of parasitoid cocoons or groups of cocoons, called clusters. They collected over 1200 gregarious clusters. They also collected over 1600 solitary cocoons.
The scientists knew if a cocoon had been parasitized based on what insect hatched out of the cocoon. Here we see a wasp hatching out of a stink bug egg. Image by US Department of Agriculture.
They raised the cocoons so they could observe what came out of each one. If a parasitoid came out, they cocoon hadn't been attacked by a hyperparasitoid. But if a hyperparasitoid came out, it's because a hyperparasitoid had laid eggs inside the cocoon. This let them test the percentage of parasitoid cocoons that are attacked in the field. They were also able to figure out which species of hyperparasitoid attacked the cocoons.
Clusters were more likely than solitary cocoons to have at least one hyperparasitoid come out. The clusters also seemed to be a hot spot for hosting all kinds of wasp hyperparasitoids. Over 17% of the clusters had been attacked by more than one hyperparasitoid species.
Clustered cocoons were more likely to be parasitized than solitary cocoons. Image by Hans Hillewaert.
Some clusters of gregarious cocoons were never attacked. But, if a cluster was attacked by hyperparasitoids, they laid eggs in the majority of the cocoons. In these attacked clusters, 65 to 81% of the cocoons held hyperparasitoid wasps.
Over three years of collection, an average of 20 to 55% of gregarious cocoons were attacked by wasps. Solitary cocoons were attacked at a much lower rate—5 to 15%. This shows that hyperparasitoids can more easily find the gregarious cocoon clusters. It also shows that the hyperparasitoids take advantage of the clusters. They laid eggs in many of the cocoons rather than looking for new clusters.
Click to enlarge.
Figure 7. Relative abundance of primary parasitoids and hyperparasitoids from primary parasitoid cocoons in the field.
Primary and hyperparasitoid wasps were reared from cocoons of the gregarious Cotesia glomerata (left) and solitary C. rubecula (right) that had been collected from Brassica oleracea during three field seasons. Colors indicate the different parasitoid species; the white segment of the bars depicts the primary parasitoids (a) C. glomerata and (b) C. rubecula; colored bar segments represent the most abundant hyperparasitoids: Acrolyta nens (1, blue bar), Lysibia nana (2, orange bar), Pteromalus semotus (3, yellow bar),Mesochorus gemellus (4, red bar), and Baryscapus galactopus (5, green bar). Photograph credit: Tibor Bukovinszky.
doi:10.1371/journal.pbio.1001435.g007
This figure shows the species of wasp that came out of parasitoid cocoons from the field. The gregarious cocoons are shown on the left and the solitary on the right.
Overall, hyperparasitoids preferred gregarious cocoons, but this varied by species. The species on top attacked more gregarious cocoons while the species on the bottom attacked more solitary cocoons.
The scale on the bottom shows the relative amount of all the parasitoid cocoons that were collected. On the left, the entire bar (white and colored) shows all the gregarious cocoons collected in a single year. Relative amount means these are shown as a percentage. So even though there were over 1200 gregarious clusters collected, these are shown as 100 percent.
Within the full bar, each color shows the percent of cocoons attacked by different species of wasp. The hyperparasitoid wasp represented by the orange bar laid eggs in a lot of gregarious cocoons in 2005 and 2007. The wasp represented by the red bar laid eggs in most of the attacked solitary cocoons in 2005 and 2007.
Click to enlarge.
Table 4. The effect of Cotesia species and year on the fraction of hyperparasitism in field experiments carried out in 3 consecutive years.
doi:10.1371/journal.pbio.1001435.t004
This table shows that hyperparasitism rates changed with species and with year. Both P values above are <0.001, which is very significant (remember, if it's below 0.05, it's significant).
Click to enlarge.
Table 5. Hyperparasitoid species and the number of hyperparasitoid wasps emerging from Cotesia glomerata and C. rubecula cocoons collected during a 3-year survey.
i:10.1371/journal.pbio.1001435.t005
This table shows the number of cocoons attacked by hyperparasitoid wasps. The wasps are listed by species. The number of cocoons are listed for both gregarious and solitary parasitoid wasps.
The discussion is usually close to the end of the paper, but it is sometimes the most interesting part.
This is where scientists talk about what they found and what it means. They will compare their data to other studies. Finally, they will come to some conclusions about what they observed. Remember, these aren't necessarily the only conclusions possible from the data. Maybe you can try to think of additional ways to interpret the data.
If the scientists had any problems during the experiment, this is also where they will discuss those.
Our results show that hyperparasitoids use plant volatiles to locate cocoons of their parasitoid host and that a network of interactions between the parasitoid, herbivore, and its food plant is involved in providing hyperparasitoids with cues of host presence. Interestingly, only one of the two parasitoids studied here altered the response of its herbivorous host with the food plant and thereby gave away its presence to hyperparasitoids. This gregarious parasitoid species, C. glomerata, was most frequently attacked by hyperparasitoids in the field, indicating that plant volatiles differentially expose parasitoid species to their enemies.
Often in science, you have to do a lot of different tests to figure out the answer to one main question. Usually you are trying to discount incorrect answers until you are left with one correct answer. These experiments led to one main finding. Hyperparasitoids can use plant chemicals to find host cocoons.
The interaction between levels of this food web changes with species.
But plants, caterpillars, parasitoids, and hyperparasitoids don't always interact the same way. The interaction can change with species. Only the gregarious parasitoid altered the plant chemicals. It did this by changing the chemicals in caterpillar throw up. Those changes caused it to be attacked more than the other species.
This change shows us something very important. A plant can change the relationship between some parasitoids and hyperparasitoids.
The two primary parasitoids that are suitable hosts for the hyperparasitoid L. nana largely differ in how they interact with their herbivorous host. C. glomerata larvae dynamically regulate the growth of their host in accordance with brood size and the concomitant amount of resources necessary to maximize adult parasitoid body size [26]–[28]. In contrast, C. rubecula–parasitized caterpillars are developmentally arrested in the third or fourth instar, depending on instar parasitized, and consume much less plant tissue than healthy caterpillars [19],[29]. However, the amount of feeding damage per se did not explain that L. nana discriminated C. glomerata–parasitized from unparasitized caterpillar-damaged plants. Controlling for the amount of damage and application of oral secretions of caterpillars revealed that changes in the oral secretions underlie the effect on differential plant responses to parasitized and healthy caterpillars. The oral secretions of parasitized caterpillars can be visually distinguished in terms of color from oral secretions of healthy caterpillars, and several parasitoid species have different effects on the color of oral secretions [20]. Clearly, the oral secretions differ in composition among caterpillars parasitized by different parasitoids, and the nature of those changes merits further investigation. Nevertheless, application of oral secretions of unparasitized or parasitized caterpillars to plant wounds has been found to result in differential expression of genes underlying the herbivore-induced plant volatile emission [20]. Here, we show that differences in induced responses of plants to caterpillars in which different parasitoids develop results in different blends of volatiles produced by the plant. However, these induced changes in volatiles were only present when C. glomerata had parasitized the herbivore and not when C. rubecula parasitized the same herbivore species. This may be caused or confounded by the lower amount of damage that C. rubecula–parasitized caterpillars make when compared to C. glomerata–parasitized caterpillars. Alternatively, it may suggest that C. rubecula has evolved to reduce its conspicuousness by not affecting the elicitors in oral secretions of its host that may reveal its presence to its hyperparasitoid enemies, whereas C. glomerata does reveal itself through such changes. In its introduced range in the United States,C. rubecula is frequently hyperparasitized [21]. This may suggest that in its invasive range C. rubecula has not (yet) adapted to be inconspicuous when developing in P. rapae, because of the shorter co-evolutionary relationship between P. rapae and C. rubecula in the invasive range. Because of the differences in attraction of hyperparasitoids to volatiles emitted by plants induced with caterpillars parasitized by different parasitoid species, we hypothesize that plant responses to herbivory result in differential selection pressure of hyperparasitoids on primary parasitoids. The observation of variation in the attraction of hyperparasitoids to caterpillars parasitized by different parasitoids that are suitable hosts to the hyperparasitoid also raises the question of whether hyperparasitoids may be able to use plant odours to assess whether a herbivore is parasitized by a nonsuitable parasitoid host. This area certainly merits further investigation.
The two parasitoid species interact with the caterpillar differently. The gregarious parasitoids balance their own growth with that of their host. This allows the host to keep growing. But the solitary parasitoids stop the growth of their host. These parasitized caterpillars eat much less than normal.
The oral secretions of some caterpillars affect the chemicals released by the plants they eat. Image by J.M. Garg.
But the scientists learned the amount of feeding didn't cause the plant to release different chemicals. Chemicals were changed by the differences between the throw up (oral secretions) of the caterpillars.
The oral secretions of parasitized and healthy caterpillars are very different. Even the color is different. And these secretions, when put on plant wounds, can cause the release of different chemicals. This might seem strange. How does that work? The chemicals in the oral secretions change the way the plant's genes work. This then causes changes in the chemicals they release.
These experiments show that host caterpillars can change which chemicals plants release. But this didn't happen for both species of parasitoid host. The gregarious wasp changed the plant chemicals, but the solitary wasp did not.
Why didn't the solitary wasp have the same effect? At first scientists thought it might be because they do less damage to the plants. Or, not changing the plant's chemicals may help the solitary parasitoid avoid being found by hyperparasitoids. This may be an adaptation.
Do gregarious parasitoids attract hyperparasitoids because the caterpillars they infect damage plants more? Or do the solitary wasps avoid detection because of an adaptation? Image by Beatriz Moisset.
The scientists also say that solitary parasitoids are spreading into new parts of the United States. This allows them to compare animals in the new parts of the range to those in old parts of the range. In the new areas of its range, the species is being hyperparasitized more often. This suggests that in the old part of the range, the wasps may have different adaptations that help them avoid being found.
If this is true, the scientists thought that in the new areas of the range, caterpillars would be different. There, they may still have changing oral secretions. This would cause increased rates of parasitism. That is exactly what other scientists are observing.
Hyperparasitoids are attracted to some plant chemicals but not others. Because of this, there is room for the parasitoids to adapt to avoid hyperparasitism. A lot of questions remained unanswered though. In particular, the scientists are curious if wasps can not only use chemicals to find hosts, but to stay away from other hosts. This may help them stick to only those hosts that will help them produce healthy young wasps.
Fitness Consequences of Response to Delayed Rewards
Secondary hyperparasitoids that are attracted to volatiles emitted by plants that are damaged by parasitized caterpillars containing fully grown parasitoid larvae may suffer fitness costs by arriving too early on a plant (i.e., before the larvae of the their primary parasitoid hosts have emerged and constructed cocoons). However, pupae themselves do not interact with plants, and plants on which no active feeding takes place are likely to have reduced volatile emission and are, therefore, more difficult to detect [1]. In addition, hyperparasitoids are even further constrained in host searching as they can only successfully parasitize young pupae of primary parasitoids (i.e., within several days after they are formed) [30]. Therefore, hyperparasitoids are likely to have evolved to respond to highly detectable and reliable cues that predict the presence of available pupae in the near future and require hyperparasitoids to wait, rather than responding to cues that may result in arriving too late.
Similar waiting strategies have been observed for pupal parasitoids that are similarly constrained in terms of host suitability (i.e., they can only parasitize the pupae of herbivores shortly after these are formed). We have observed on numerous occasions that the pupal parasitoid Pteromalus puparum sits on or next to final instar Pieris caterpillars awaiting their pupation (E.H. Poelman and J.A. Harvey, unpublished observations). For both hyperparasitoids and pupal parasitoids, natural selection may favour such a waiting strategy even further, because healthy and parasitized caterpillars are well known to leave the host plant during the wandering phase and to climb onto a neighbouring plant and are thus not detectable through volatiles emitted by the plant on which they were previously feeding. The strategy of herbivores or parasitized herbivores to wander off the plant on which they have been feeding suggests that selection is being imposed on such behavior, which is likely to be mediated by responses of their own natural enemies such as hyperparasitoids.
Despite the potential delay of arriving too early and having to wait on the plant to be able to parasitize the primary parasitoid pupae, the plant volatile cues derived from feeding by parasitized herbivores are the most detectable cues predicting host presence. L. nana females generally carry up to 40 mature eggs after several days that are ready for oviposition and thus can exploit an entire brood of C. glomerata within several hours [30]. Moreover, a female can mature an additional 20–30 eggs over the course of 24 h [30]. This suggests that L. nana has probably co-evolved with host species such as C. glomerata because of the strong synchrony between egg load dynamics in the hyperparasitoid and average cocoon cluster size in C. glomerata [31].
We've heard it said before: timing is everything. This is also true for wasps. If hyperparasitoids want to find their parasitoid host in a cocoon, they must find them at just the right time.
Wasps that find their hosts too early may have to wait for parasitoid larvae to form cocoons. Image by Alvesgaspar.
Parasitoids form cocoons on the outside of the caterpillar. These cocoons are where the hyperparasitoid lays its eggs. If the hyperparasitoids find the caterpillar too long before the parasitoid cocoons form, they have to wait around to lay their eggs.
But too late is even worse. Hyperparasitoids can only lay in cocoons that have formed within a few days. And, if they find the cocoon after that point, the hyperparasitoid won't be able to lay eggs in those hosts at all. So it's most likely that they use signals that tell them when a host is almost ready. This lets the wasps find the host in time, without having to wait too long.
This importance of timing may be why hyperparasitoids use signals to find their host. Over time, the wasps that used such signals ended up having more young. This is how the ability would have evolved, or spread through the population. Such waiting behavior has been found in parasitoids as well.
Parasitic wasps need to respond to chemical signals quickly and arrive to find their prey early. Otherwise, they may find the caterpillar has left or their hosts may be too old. Image by
Arriving slightly early may be useful for other reasons too. A parasitized caterpillar could wander away from a plant it was eating. The plant may still be releasing chemicals that attract hyperparasitoids. However, the caterpillar is long gone. In this case, the wasps should arrive just before they need to lay eggs so they don't miss out.
In addition, some hyperparasitoids lay many eggs that allow them to use a whole group of parasitoid cocoons. The number of eggs match the number of cocoons very well. This makes the scientists think that the two have co-evolved. Co-evolution means that when characteristics of one species changes, a related characteristic in the other changes too.
Foraging Decisions on the Food Plant
Foraging decisions of the hyperparasitoids on the food plant clearly underlie the contrast between C. glomerata and C. rubecula hyperparasitism levels when cocoons of C. glomerata and C. rubecula were exposed to the natural hyperparasitoid community on plants damaged by parasitized or healthy caterpillars.
The field study in 2011 showed a preference of hyperparasitoids for plants damaged by gregariously parasitized caterpillars, and this was reflected in hyperparasitism rates on gregarious C. glomerata pupae (Figure 6), supporting our findings in the laboratory choice assays. However, the effects of the herbivory treatments in the field assay did not prevail on solitary pupae of C. rubecula. Moreover, we also found a higher hyperparasitism rate of L. nana in the solitary pupae in the field season of 2011 compared to the field seasons of 2005 to 2007. Several factors, which are not necessarily mutually exclusive, may account for this: first, our method of offering pupae on paper may expose solitary cocoons more to hyperparasitoids than occurs in nature. Second, when considering the total number of pupae in a brood, we offered more C. glomerata than C. rubecula pupae. Although more hyperparasitoids were recovered from gregarious pupae, the rates of individual clutches of C. glomerata that contained any hyperparasitoid and solitary pupae of C. rubecula that were hyperparasitized were similar. The hyperparasitism rates on C. glomerata underestimate the actual rates at which clutches were found by more than one hyperparasitoid. Third, due to the setup of this field study, we excluded hyperparasitism by primary hyperparasitoids (that oviposit in the parasitoid larvae when these develop within the caterpillars). Therefore, L. nana might encounter less competition from primary hyperparasitoids and may therefore alter its oviposition strategies. Fourth, as described above, female hyperparasitoids, such as L. nana, may exploit a large proportion of their host pupae once they locate a host clutch [31]. The hyperparasitoids locating a gregarious brood spend more time on the brood and are egg limited when exploiting the whole brood, whereas they are time limited when exploiting solitary pupae. Therefore, hyperparasitism rates may have been elevated on solitary C. rubecula pupae despite the larger number of eggs laid in gregarious broods.
Every experiment has problems. Here, the scientists discuss some things that may have changed their findings.
Hyperparasitoids use plant signals to find hosts. In the field, they preferred chemicals from plants eaten by gregarious parasitoid hosts. In the lab, this was partly true, but the results didn't all match up.
Remember our plant treatment groups?
Undamaged.
Damaged by healthy caterpillars.
Damaged by caterpillars hosting solitary parasitoids.
Damaged by caterpillars hosting gregarious parasitoids.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
When only solitary parasitoids were around, hyperparasitoids didn't prefer any of the groups. Also, preference seemed to change over time. Over the years of the study, the wasps' seemed to like solitary parasitoids more.
The scientists explain that the experimental set up may have increased parasitism beyond what was natural. In the experiment, they put the hosts on little pieces of paper. This may have made them easier to find.
The scientists also ended up with more gregarious parasitoids available than solitary ones. They used clusters, or groups of eggs, for the gregarious species. But they were able to control for this. If a whole cluster was considered equal to one solitary egg, the wasps chose the hosts equally.
One thing scientists didn't take into account is that not all parasitic wasps lay eggs in cocoons. Click for more detail.
What's more, they also underestimated the number of hyperparasitoids that laid eggs in gregarious cocoons. If two of the same species found the same clutch, the scientists couldn't tell the difference. So they only estimated one hyperparasitoid in such cases.
The scientists also did not take into account other types of hyperparasitism. Some wasps lay eggs inside the parasitoids before they come out of the caterpillar. This wasn't counted, but may have affected the number of available host cocoons.
Finally, the scientists didn't control for the different behaviors the hyperparasitoids show with each parasitoid.
This parasitoid wasp only lays one egg per pupa or cocoon. After that, she must go look for another pupa. The number of eggs she lays is limited by how long it takes her to find pupae. Image by the USDA.
When on the gregarious cocoons, the hyperparasitoids may not be able to lay as many eggs as they'd like. This makes them "egg-limited" because the number of cocoons can affect their success.
When on the solitary cocoons, the wasps only have one cocoon in which they can lay. This makes them "time-limited" because they need to lay quickly. Then they can go find other cocoons in which to lay.
The scientists present all of these potential problems so we know what may have thrown off some of their data.
Plant Fitness Benefit of Volatile Release
Our study shows that enemies of those natural enemies that benefit plant fitness may also use plant-produced odours to find their hosts or prey. In this way, the plant may be caught between a “rock and a hard place,” in that two out of three trophic levels of consumers that are detrimental to the plant (either directly, through herbivory, or indirectly, through a reduction in the abundance of beneficial carnivores at the third trophic level, caused by organisms at the fourth trophic level) benefit from using herbivore-induced plant volatiles. The beneficial effect on plant fitness of attracting parasitic wasps, to indirectly defend itself against their herbivore attackers, has been intensively discussed [2],[3],[6]. Although it has been recognized that volatiles released by plants that are under attack by herbivores provide parasitoids and predators with a cue that can be used in host location, the presence of active “signalling” and associated selection on plants that are stronger signallers has thus far received less attention. Although some studies have reported a fitness benefit of plants on which herbivores were attacked by parasitoids [10]–[12], other studies have reported negative fitness consequences of plants emitting volatiles by becoming more apparent to herbivores [8]. It is important to emphasize that volatile cues may provide many community members with information and thereby may not necessarily result in a fitness benefit to plants [6]: although plant volatiles may function as a “cue” to parasitoids, they may not be a specific “signal” released by the plant (implying a selective benefit). Although short-term negative consequences of attracting hyperparasitoids for plants may be absent, as the hyperparasitoid is not affecting the direct benefit of reduced herbivory by parasitized caterpillars, the plant may be presented with a cost of reduced population size of its beneficial natural enemies when a next generation of herbivores arrives. Our results show that hyperparasitoids may parasitize up to 55% of the parasitoid offspring, therefore potentially playing a major role in parasitoid population dynamics. Furthermore, the parasitoid species studied here have been found to parasitize over 90% of the herbivores when parasitoids are at their peak abundance during the season [32]. The effect of parasitoid–hyperparasitoid interactions therefore may have significant consequences for herbivore populations, and thereby indirectly hyperparasitoids may significantly contribute to selection on plant traits such as volatile release.
Caterpillars damage plants and parasitoids damage caterpillars. What does that mean for the plant? It means that parasitoids may benefit the plant by hurting the caterpillars.
If parasitoids hurt caterpillars, they indirectly help plants. So then if hyperparasitoids hurt parasitoids, they indirectly hurt plants. Image by Scott Bauer of the USDA.
So how do hyperparasitoids fit into this? These wasps harm parasitoids. Because parasitoids are good for the plant, this means hyperparasitoids may end up hurting the plants in a way.
The plant chemicals are another story. We know those chemicals can attract parasitoids. But, they can also attract more animals, including ones that eat the plant. This makes it unclear if releasing chemicals helps the plant.
If the chemicals don't help the plant, maybe they aren't releasing them as a signal. It is possible that parasitoids have simply learned to use the chemicals as a signal.
Overall, the effect of hyperparasitoids on plants is unclear, but a few things are likely.
Scientists are still unsure of all the effects hyperparasitoids have on cabbage and other plants. However, it's likely they have a long-term negative effect. Image by Scott Bauer of the USDA.
There probably isn't any short-term negative effect on the plant. The caterpillars have already been parasitized, potentially helping the plant.
There is likely a long-term negative effect on the plant population. Hyperparasitoids may parasitize up to 55% of parasitoid offspring. This means parasitoid population will be much smaller the next year. In that next year, fewer caterpillars will be parasitized.
Sometimes we save the best for last. If an article has a conclusion, this is where the authors summarize all the important findings from the whole study. This is the best section to read if you want a shorter description of the overall importance of the study.
The fitness consequences of the emission of herbivore-induced plant volatiles are dependent on the quantitative composition of the plant-associated community at several trophic levels. The fitness benefit of volatile release should, therefore, be evaluated in the natural context of the plant-associated insect community including fourth-trophic-level organisms [6]. This will help to improve our understanding of the function of herbivore-induced volatiles in plants and of how the ecological effects of volatiles can shape the life histories of species interacting in insect communities associated with these plants [33],[34]. Furthermore, these findings are important in the context of developing Integrated Pest Management strategies in which herbivore-induced volatiles of crops are manipulated to optimize the control of insect pests by using parasitoids. Overexpression of herbivore-induced plant volatiles in crops or field application of synthetic parasitoid attractants may not benefit pest control in conditions where the responses of hyperparasitoids to HIPVs cause major mortality to parasitoids [35].
The chemicals released by the plant can affect animals in the area. This can change the plant's success.
Scientists may be able to use the findings from this study to help design new ways to control pest species, like this brown marmorated stink bug. More studies are needed before that can happen though. Image by Stephen Ausmus of the USDA.
In order to better understand this, the scientists suggest that more studies should be done. But, they warn that those studies need to include animals from various trophic levels to see the big picture.
The findings from this study may be important to help protect crops from insect pests. If plants could be made to release certain chemicals, it could attract parasitoids. Those parasitoids may be able to help control problem insects.
But, this study shows that hyperparasitoids can also use the plant chemicals as signals. How hyperparasitoids fit into this idea of pest control needs to be studied more before it will be useful.
The Materials and Methods section helps you form a picture of exactly what the scientists did. These two scientists are on a bat collecting trip. Image by US Fish and Wildlife Service Headquarters.
The Materials and Methods section is where the scientists explain exactly how they did the experiments. This section is often the most dense and detailed. The scientists try to write it in a way that would allow someone else to repeat the experiment in exactly the same way.
It’s good for the scientists to explain everything they did. But sometimes, they use certain words or methods that even other scientists don’t understand.
We even had to check with a few scientists to make sure we understood this section correctly. Don’t worry if you have problems understanding it. Just give it a shot. Our explanations are included to help you through it.
Plants and Insects
Brassica oleracea var gemmifera cv. Cyrus plants used for olfactometer experiments were grown in 1.45-l pots containing peat soil (Lentse potgrond, no. 4, Lent, the Netherlands) and provided with SON-T light (500 µmol/m2/s; L16:D8) in addition to natural daylight in a glasshouse compartment (18–26°C, 50%–70% r.h.). When plants were 4 wk old, they were fertilized weekly by applying 100 ml nutrient solution of 2.5 mg/l Kristalon Blauw (Hydro Agri Rotterdam, the Netherlands (N-P-K-Mg) 19-6-20-3) to the soil and used in experiments when they were 7 wk old. To prepare parasitized caterpillars for the induction treatments, individual first instar P. rapae larvae were exposed to a single female C. glomerata or C. rubecula, which were allowed to parasitize the caterpillar in a glass vial. For C. glomerata, caterpillars were considered to be parasitized when the wasp had inserted her ovipositor in the caterpillar for at least 5 s. For C. rubecula, because of herbivore immune responses to parasitoid eggs [34], the wasp was allowed to oviposit 3 times in the same caterpillar, to increase the success rate of parasitism. Due to larval cannibalism among the parasitoids, only a single C. rubecula larva would develop eventually [36]. The hyperparasitoid L. nana was reared on C. glomerata cocoons in the absence of plant and herbivore-derived cues.
Green or glasshouses are structure frames that are covered with plastic or glass. Click for more detail.
In this first section of the methods, we learn where the scientists got the plants and the insects for this study. Plants were grown in pots and were kept in a greenhouse (called a glasshouse). They were fertilized as they grew and used for the experiments when they were seven weeks old.
To allow caterpillars to be parasitized, they were put in a glass vial with a parasitoid wasp. This was done when the caterpillars were in an early stage of growth, the first instar larvae.
How did the scientists make sure that the caterpillars were parasitized? This method varied by species.
Scientists made sure caterpillars were parasitized by letting parasitoids insert their ovipositor. Click for more detail.
Gregarious parasitoid wasps had to insert their ovipositor into the caterpillar for at least five seconds. Ovipositors are the long, piercing body part that these wasps use to lay eggs.
The solitary parasitoids were allowed to insert their ovipositor three times in the same caterpillar. They should lay one egg with each insertion. This way, at least one of the eggs would develop into a larva. If more than one developed, the larvae would compete against each other. In the end, only one would end up living inside the caterpillar.
The scientists raised hyperparasitoid wasps in gregarious parasitoid cocoons.
Y-Tube Olfactometer Assays
All Brassica oleracea var gemmifera cv. Cyrus plants for the olfactometer assays were treated 24 h before the tests. First, plants were infested with either two unparasitized fourth instarPieris rapae caterpillars or two fourth instar caterpillars that contained fully grown parasitoid larvae of either C. glomerata or C. rubecula as a result of parasitization of the caterpillar in their first instar. In a second experiment with oral secretions of caterpillars, plants were artificially damaged with a pattern wheel by drawing three lines of 3 cm long on each of the four youngest fully expanded leaves and treated with 25 µl of caterpillar oral secretions onto the damaged sites. Oral secretions were collected from healthy and C. glomerata–parasitized fourth instar P. rapae caterpillars, using 5 µl capillaries. Single caterpillars regurgitated 2–8 µl that we pooled to be used in the induction treatments. We decided not to test the relative attractiveness of plants induced with oral secretions of C. rubecula–parasitized caterpillars, because it would lack biological relevance as the data from our choice assays with actual feeding damage indicate that the quantity of damage by gregariously parasitized caterpillars is likely to explain the preference of hyperparasitoids for treatments with higher amounts of leaf damage (Figure 4). We have restricted to testing the effect of qualitative differences in the oral secretion of parasitized and unparasitized caterpillars only to treatment combinations where we did not identify a statistical difference in the amount of damage between the treatments.
This section details the wasp choice tests. Plants were damaged 24 hours before they were exposed to hyperparasitoids.
For these two experiments, wasps had to choose between plants based only on caterpillar damage. No cocoons were present. Click for more detail.
In the first experiment, plant treatments were natural. Healthy or parasitized caterpillars were placed on specific plants. Once the plants were damaged, they were used in the wasp choice test. Hyperparasitoids were tested to see if they preferred plants damaged by certain caterpillar hosts.
In the second experiment, the scientists damaged the plants. They did this to control the amount of damage and caterpillar presence. Then, they put caterpillar oral secretions (throw up) on the plant wounds. This let the scientists compare the effects of throw up from both healthy and host caterpillars. The parasitized caterpillars were all hosting gregarious parasitoids.
Scientists used capillary tubes to collect oral secretions from caterpillars. Then they rubbed them on plant wounds before the wasp choice test. Image by GOKLuLe
How do you think you'd go about trying to collect caterpillar throw up? The scientists did it using tiny glass tubes called capillary tubes. If held at the right angle, the small tubes suck up liquids, caterpillar throw up included.
The scientists only used throw up from healthy and gregarious host caterpillars. Wasp preference for gregarious parasitoids hosts was likely due to the amount of damage those caterpillars caused. By leaving out solitary parasitoid hosts, the scientists could focus their question. They tested two main things with this experiment. First, they tested if parasitism changed caterpillar throw up. Further, they tested if those changes affected the plant chemicals that the hyperparasitoids used as a signal.
To test whether volatiles derived from oral secretion itself may be attractive to hyperparasitoids, we applied 25 µl oral secretion of C. glomerata–parasitized P. rapae caterpillars onto undamaged plants with a fine brush. We tested the relative attractiveness of the oral-secretion-treated plants to undamaged plants treated with 25 µl of water.
To test if it was just caterpillar throw up or the effect of it on plant wounds, scientists tested if wasps were more attracted to throw up or water on healthy plants. Image by Frank Vincentz.
The scientists also wanted to know if the caterpillar throw up alone would attract hyperparasitoids.
They painted a little of the caterpillar throw up onto undamaged plants. They also painted water onto undamaged plants.
The water was a control, to test if the hyperparasitoids were attracted to undamaged plants for some other reason.
Shortly before L. nana females were tested for their behavioral response to plant volatiles in Y-tube olfactometer bio-assays, we removed caterpillars and their feces from the plants and placed the plants in one of two glass jars (30 l each) that were connected to the two olfactometer arms. A charcoal-filtered airflow (4 l/min) was led through each arm of the Y-tube olfactometer system, and a single wasp was released at the base of the stem section (3.5 cm diameter, 22 cm length) in each test [37]. Wasps that passed a set line at the end of one of the olfactometer arms within 10 min and stayed there for at least 15 s were considered to have chosen for the odour source connected to that olfactometer arm. To compensate for unforeseen asymmetry in the setup, we swapped the jars containing the plants after testing five wasps and replaced the set of plants by a new set of plants after testing 10 wasps. The Y-tube olfactometer setup was placed in a climatized room, and in addition to daylight it was illuminated with four fluorescent tube lights (FTD 32 W/84 HF, Pope, the Netherlands).
Now we're going to jump back to the first experiment for a minute. After the caterpillars were done damaging the plants, the scientists took them off the plants. They picked off any caterpillar poop that was left on the plants (in case this had any effect).
For the choice tests, wasps were released into a Y-maze. Click for more detail.
The plants were then used in a choice experiment. Two plants (from different treatments) were placed into two glass jars. The jars were attached to a small series of pipes in the shape of a Y. The top of the Y connected to the two jars and one by one, wasps were released into the base of the Y.
Clean air was pushed through the jars and into the two arms of the Y, giving the wasps a choice for which plant had a smell that they preferred. When wasps spent more than 15 seconds near one plant, that plant was counted as the choice of the wasp.
Plants were switched after five tests to control for the side of the Y maze in which they were placed. Additionally, plants were only used for 10 tests, after which new plants were used. The tests were run in a temperature-controlled room that had both lamp light and natural (sun) light.
Field Assay Hyperparasitoid Attraction to Plant Volatiles
Eighty-four-week-old plants were transplanted into the field with 1×1 m spacing between plants and allowed to adjust to field conditions for 1 wk. Thereafter, the plants were subjected to one of four induction treatments: (1) not treated with herbivory (i.e., undamaged controls, UD), (2) infested individually with either two unparasitized first instar P. rapae caterpillars (PR), (3) two C. glomerata–parasitized P. rapae caterpillars (PR-CG), or (4) two C. rubecula–parasitized P. rapae caterpillars (PR-CR).
Unparasitized and parasitized caterpillars were allowed to feed on plants for 10 d, which was approximately the whole development period of the Cotesia larvae. Each plant was covered with a fine-mesh net when planted to avoid other herbivore infestations on the foliage and to prevent the herbivores used for induction to wander off the plant.
To test the effects of plant induction with different types of herbivory on hyperparasitism, we attached parasitoid pupae onto the plants in the field. Individual pupae of C. rubecula, or clutches of C. glomerata, were first attached to a paper disc (3×3 cm) with a small droplet of glue. We removed nets and caterpillars just before attaching the paper discs carrying the pupae with a pin needle. Half of the plants for each treatment received five C. glomerata clutches, and the other half received five C. rubecula pupae. The pupae were exposed to the natural community of hyperparasitoids and recollected after 5 d. They were kept separately in 2.2 ml Eppendorf tubes that were closed with cotton wool. The Eppendorf tubes were checked daily for emerging primary parasitoids and hyperparasitoids. All wasps were identified to species level. A completely randomized design was applied to the field studies. Five replications, each with 80 plants (10 replicates of each treatment), were carried out from June until October 2011.
Even in field experiments, it is good to try to control as many variables as possible. Scientists planted plants of the same species and age to use in the field. Plants were spread apart evenly and grew in the field for one week before the scientists gave them one of four treatments.
We've seen all these plant treatments before, but just as a reminder, they were:
Undamaged.
Damaged by healthy caterpillars.
Damaged by caterpillars hosting solitary parasitoids.
Damaged by caterpillars hosting gregarious parasitoids.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
For a week before treatments and during treatments, the plants were covered with nets. This kept other insects from damaging them and kept the caterpillars the scientists placed on the plants from wandering away. Caterpillars fed on the plants for 10 days.
Scientists wanted to see if the hyperparasitoids liked a certain combination of treatments. They paired plants from the different treatments with cocoons of the two parasitoids. There would be eight combinations in all.
Within each plant treatment, half the plants were paired with the solitary parasitoid (top) and half with the gregarious one (bottom).
To do this, the scientists put cocoons on the plants. In each treatment, half of the plants had gregarious parasitoid cocoons and half had solitary ones. They attached five groups of the chosen cocoon type to each plant.
How do scientists attach cocoons to plants? With glue, paper, and needles, of course. The scientists glued a small, circular piece of paper to the cocoon. Then they pinned these papers to plants using tiny needles.
After the cocoons were attached to the plants, the scientists left them out with the natural predators in that area for five days.
To allow local hyperparasitoids to access the cocoons, the paired treatments were grown outside and uncovered for five days before the cocoons were collected. Image by sweetbay103.blogspot.com.
To see if and by what species a cocoon was parasitized, scientists then isolated the cocoons. Groups of gregarious cocoons were kept together, but solitary ones were each held in their own container.
Containers were checked daily to see which species of hyperparasitoid had laid eggs in the cocoons.
As in all good experiments, the scientists redid, or replicated, the test many times. This helped to make sure they had enough samples, or a large sample size. Eighty plants were used for each full round of the experiment (10 plants in each of 8 treatment combinations). The scientists redid this part of the experiment five times to make sure they had enough samples.
Field Collections of Parasitoid Cocoons
To assess hyperparasitism rates and species communities on the solitary and gregarious primary parasitoid, we established plots of 6×6 m containing 49 plants of B. oleracea cultivars in an experimental field in the vicinity of Wageningen, the Netherlands, during 3 consecutive years (2005–2007). Within plots, plants were planted in a square of 7×7 plants with a spacing of 75 cm between plants. Plots were isolated by strips of 6 m wide that were sown with a grass mixture of Lolium and Poa species. During the growth season of cabbage plants, from early May until the end of September, we conducted weekly surveys on the plants for C. glomerataand C. rubecula pupae by investigating both sides of all leaves of all plants in the plots. The pupae were collected and placed individually in 2.2 ml Eppendorf tubes and closed with cotton wool. The pupae with their external silk cocoon were weighed, and for the gregarious C. glomerata, the brood size was determined. The Eppendorf tubes were checked daily for emerging parasitoids, which were individually transferred to another Eppendorf tube and stored at −20°C. All wasps were identified to species level.
To collect parasitoid cocoons, scientists maintained plots of cabbage and monitored them throughout the cabbage growing season. Their plots were not as dense as the one shown here. Modified image from CIAT.
In the field experiment, the scientists used plants that they had grown in lab. By doing this, they could make sure they were all the same age and make sure they were evenly spaced outside. Each week during the field season (May until September), they checked each leaf of each plant for cocoons.
When found, parasitoid cocoons were collected and kept in small storage tubes. The scientists measured the number of cocoons and how much they weighed. Any parasitoids that came out of the cocoons were identified.
Plant Volatile Analysis
Headspace collection of plant volatiles.
To characterize the differences in plant volatile release after herbivory by unparasitized and parasitized caterpillars, we collected the headspace of 6-wk-old B. oleracea plants subjected to different herbivore treatments. Dynamic headspace sampling was carried out in a climate room, and we collected 12 replicates of each of four experimental treatments: (1) undamaged plants, plants infested with two P. rapae caterpillars that were either (2) unparasitized or parasitized by (3) C. glomerata or (4) C. rubecula. Caterpillars were allowed to feed for 7 d, during which the parasitoid larvae nearly completed their development inside the herbivore. Prior to volatile collection, caterpillars were removed and plants were removed from their pots. Their roots and soil were carefully wrapped with aluminium foil. During volatile collection, the plants were placed individually into a 25 litre glass jar, which was sealed with a viton-lined glass lid with an inlet and outlet. Compressed air was filtered by passing through charcoal before reaching to the glass jar containing the plant. Volatiles were trapped by sucking air out of the glass jar at a rate of 150 ml min−1 through a stainless steel tube filled with 200 mg Tenax TA (Markes, Llantrisant, UK) for 4 h.
Hyperparasitoids most liked plants that were damaged by caterpillars hosting gregarious wasps. But which plant chemicals were the wasps using to find the caterpillars?
To test this, scientists collected the "headspace" of plants. Basically, headspace is the air that closely surrounds the plant. The scientists collected this air from the headspace of plants from our same four plant treatments:
Undamaged.
Damaged by healthy caterpillars.
Damaged by caterpillars hosting solitary parasitoids.
Damaged by caterpillars hosting gregarious parasitoids.
Examples of the four plant treatments. From left to right, images by Griensteidl, James K. Lindsey, Scott Bauer of the USDA, and Stsmith.
Twelve plants were in each treatment.
In the treatments with caterpillars, the caterpillars fed on the plants for seven days. They were removed from the plants before the air was collected.
To collect headspace, plants are enclosed in a mostly sealed container with one way for air to come in and one way for air to go out.
Collecting the headspace of a plant is a little harder than it sounds. First, the plants were taken out of their pots and the soil holding their roots was wrapped in aluminum foil, like you use to cover leftover food.
The plants with wrapped bases were then put into a glass jar that was sealed, mostly. The jar had a hole where air could enter and one where air could exit. Clean air was pushed into the jar at one end. Air was then sucked out from the other end of the jar. This was done over a period of four hours to sample the chemicals the plant released.
Analysis of plant volatiles.
A Thermo Trace GC Ultra coupled with Thermo Trace DSQ quadrupole mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA) was used for separation and detection of plant volatiles. Prior to releasing the volatiles into the GC, the Tenax TA cartridges were dry-purged under a stream of nitrogen (20 ml min−1) for 10 min at ambient temperature in order to remove moisture and oxygen. The collected volatiles were released from the Tenax TA thermally on an Ultra 50:50 thermal desorption unit (Markes, Llantrisant, UK) at 250°C for 10 min under a helium flow of 20 ml min−1, while re-collecting the volatiles in a thermally cooled universal solvent trap at 10°C using Unity (Markes, Llantrisant, UK). Once the desorption process was completed, volatile compounds were released from the cold trap by ballistic heating at a fast rate (40°C s−1) to 280°C and was then kept at 280°C for 10 min, while the volatiles transferred to a ZB-5MSi analytical column (30 m×0.25 mm I.D. ×1.00 µm F.T.; Phenomenex, Torrance, CA, USA) in a splitless mode for further separation. The column was operated at an initial temperature of 40°C and the temperature was raised at 5°C min−1 to 280°C and held for 4 min under a column flow of 1 ml min−1 in a constant flow mode. The DSQ mass spectrometer (MS) was operated in a scan mode with a mass range of 35–350 amu at 5.38 scans s−1, and ionization was performed in EI mode at 70 eV. The MS transfer line and ion source were set at 275 and 250°C, respectively. Compound identification was based on comparison of mass spectra with those in the NIST 2005 and Wageningen Mass Spectral Database of Natural Products MS libraries. Experimentally obtained linear retention indices (LRI) were also used as additional measures for confirming the identity of compounds. Relative quantification by peak areas of individual compounds was done using the integrated absolute signal of a quantifier ion in the selected ion monitoring (SIM) mode. The individual peak areas of each compound were computed into peak area per gram shoot biomass to correct for differences in size of individual plants.
Let’s think about air for a minute. The air around you and in your lungs is made up of a mixture of gases. These gases are each their own element or type of atom (nitrogen, oxygen, argon, carbon dioxide, etc.). Each atom type has its own electrical charge and its own weight. So how might you figure out what different gases are in a mixture?
Scientists can measure what gases are in a mixture by separating them into groups based on the weight and electrical charge of atoms. This is what the scientists describe above. They took gases that the plants released and ran them through a mass spectrometer (mass spec, for short) to identify the chemicals within.
The mixed gases are separated and measured using a special thermal mass spectrometer. Click to enlarge.
Mass specs use electric or magnetic fields to pull different atom types apart. But just that separation isn’t enough. Before the different chemicals can be analyzed, two important things have to happen.
First, the chemicals need to be separated as much as possible. Temperature is a big help in this process. Each chemical handles temperatures a little differently. One of the major examples of this is the temperature at which different chemicals become gas.
Second, the molecules need to get an extra boost of electrical charge. This helps them respond more strongly to the electric or magnetic fields. The boost can be given by either adding or removing electrons. (The number of electrons in a molecule changes its electrical charge.) Adding or removing electrons is called ionization.
For ionization, special gases that either take up or lose electrons easily are put into the gas mix. This encourages the transfer of electrical charge from one molecule to another.
So, now that we know the major processes that the scientists used to analyze the samples, let’s take a step back. We will go back to the beginning of this section and walk through it a bit more carefully.
Extreme heating and cooling is important to the process of separating chemicals. Flame image by Søren Wedel Nielsen. Liquid nitrogen by Robin Müller.
Let’s jump back to our plant chemicals that are waiting patiently. First, the scientists needed to remove any water or oxygen from the plant chemicals. They did this using an air stream of nitrogen that they flowed over the chemicals, drying them out and pushing out any oxygen. This would create a dry mix of the chemicals.
Here is where temperature becomes really important. This dry mix, at the right temperatures, can still be turned back into gas. The mix was heated up to 250ºC and mixed with helium, getting rid of the nitrogen. Then, the new mixture was cooled again so that the chemicals would condense out (helium doesn’t condense, as it is almost always a gas).
The chemicals trapped in this cold condensation were then turned back into gas by heating them up very quickly (40ºC per second!). This is like going from laying in the cold snow to standing in the hot desert in one tiny second. The chemicals were kept at a high temperature (280ºC) for ten minutes to make sure all the chemicals became gas.
Finally, these gases were ready to be fully separated. They were transferred into a long tube called a column, where they cooled down a bit. Starting at 40ºC—our desert temperature—the temperature was raised slowly. This caused different chemicals to become gas at different times.
Gases were run through a column where they were first cooled and then slowly heated to separate different chemicals. The separated chemicals were then analyzed in the mass spec.
These chemicals then passed through the mass spec while being ionized. The electrical fields in the mass spec started to pull atoms differently depending on mass and charge. As the atoms were pulled apart, the machine started analyzing the gas. The different chemical elements were identified based on the weight and the charge of the atoms.
From this process, the mass spec spits out a bunch of numbers. These numbers can then be compared to a database that identifies the mass and charge of all kinds of chemicals. But mass and charge aren't the only ways of identifying the chemicals. The timing of chemical release from the column was also used. This let the scientists double-check their findings, as the chemicals were released at different times due to the slowly rising temperature.
When the mass spec produces a reading, it doesn't just measure the mass and charge of the atoms. It also creates a peak based on how much of that atom is present. Using this, the scientists could measure the relative amount of each chemical that was in each sample.
The mass spec produces a reading that often looks like this. Click for more detail.
Phew! We just covered a lot of techniques and a lot of information. Using all of this data and knowledge, the scientists learned which and how much of each chemical was being released by each plant.
Statistical Analysis
L. nana preferences for herbivore-induced plant volatiles, as tested in two-choice Y-tube olfactometer assays, were analysed using two-tailed binomial tests.
Most statistical tests are used to compare sets of numbers that come in a range. For example, imagine you are comparing the weights of 30 people that exercise to 30 people that don't. There would probably be some overlap of numbers, since people vary a lot in weight. Generally, though you would expect the people that exercise to weigh less on average.
When flipping a coin there are only two choices: heads or tails. In the wasp choice experiment, there were also only two possible choices.
However, the tests that you can use change a little bit when you don't have a range of numbers. For example, in this experiment, when looking at the choice that a wasp makes, it can only choose one of two options.
It's kind of like flipping a coin... it can only be either heads or tails. In cases like this, scientists use a special test called a binomial (which means two-name or two-term) test to see if one choice (or in this case, side of the Y) shows up more often than the other.
Hyperparasitoid preferences for plant volatiles induced by unparasitized P. rapae caterpillars and caterpillars parasitized by gregarious or solitary primary parasitoids under field conditions were analysed using two Generalized Linear Models (GLMs). To analyze the effects of plant inductions with different types of herbivory on hyperparasitism at plant level, we modelled the dependent variable as a binomial occurrence of hyperparasitism per plant (400 plants equally divided over five replicates) and scored presence of hyperparasitoids in pupae as 1 and absence as 0. Additionally, to test the effects of the plant inductions on hyperparasitism at cocoon level, we modelled the dependent variable as the number of pupae or clutches giving any hyperparasitoid out of the fixed totals of five pupae attached to the plant. Into the two models we included the fixed factors caterpillar induction (undamaged, unparasitized P. rapae, P. rapae parasitized by C. glomerata, and P. rapae parasitized by C. rubecula), replicate (five replications), types of pupae (gregarious or solitary), and the interactions between the three terms.
The scientists had lots of data on hyperparasitoid preference, but how did they use that data? They had to run statistical tests. They tested to see if hyperparasitoids prefer the chemicals that plants release when they are under attack from a certain caterpillar host. For this test, they used a Generalized Linear Model.
Height, weight, and grades often follow this bell curve shape, called a normal distribution. Image by 35Abdul.
This type of statistic allows you to draw a relationship between two factors that aren’t normal. What we mean by normal here is a normal distribution—a bell curve, like you see in factors like height, weight, and grades in most classes.
Non-normal factors here include the amount of hyperparasitism or the laying of hyperparasitoid eggs on plants. When there is no regular bell curve, a Generalized Linear Model still lets you figure out how the two factors should relate.
The scientists used this statistic to test two things:
If the type of caterpillar host changed the number of hyperparasitoids that laid eggs in the caterpillars on a plant.
If the type of plant treatment changed the relative amount of eggs laid on a plant. (So, number of total cocoons versus number of cocoons with hyperparasitoid eggs for each treatment group.)
If scientists redid this experiment, they could replicate the species used as well as the plant treatments. This makes both of these fixed factors. Image by Sam Fraser Smith.
The scientists also mention fixed factors. Let's think about these factors a little differently. If the scientists redid this experiment, fixed factors could be replicated to be exactly the same. For example, here, there are four plant treatments and they could all be repeated. The number of replicates and the species used were also fixed factors.
Random factors are different. They cannot be replicated. For example, if the scientists redid the experiment, the caterpillars would all have to be new ones. They couldn’t repeat the experiment on the same caterpillars at the same age.
The final measure that the scientists discuss here is the interaction between effects. When factors interact, that means that one finding might affect another. This is an important thing to take into account when looking at the results of an experiment.
For the field collections of solitary and gregarious pupae (n = 1,668 and 1,256, respectively), we analysed whether the gregarious broods of primary parasitoids were more frequently found by hyperparasitoids than solitary pupae and whether this results in differences in the total fraction of hyperparasitism of primary parasitoid offspring from gregarious and solitary species. First, we tested whether solitary and gregarious pupae of the primary parasitoids differed in the proportion of occasions that these were found by a hyperparasitoid. For each solitary parasitoid, we scored a 1 when there was a hyperparasitoid emerging and a 0 when the primary parasitoid emerged. Because gregarious broods could be hyperparasitized to different degrees (percentage pupae parasitized), which may be a result of a single hyperparasitoid finding the gregarious brood and parasitizing several pupae or a result from multiple occasions on which hyperparasitoids found the gregarious brood, we scored a 1 when any hyperparasitoid emerged from the gregarious Cotesia brood. We used a Generalized Linear Model (GLM) to test for the effect of Cotesia species and year as well as their interaction on the binomially distributed occurrence of hyperparasitism.
Even though each gregarious cocoon is much smaller than a solitary cocoon, a cluster of them are much larger than the solitary cocoons.
For the field experiment, the scientists wanted to see if the hyperparasitoids found one species of parasitoid more often. They were also curious if this had to do with the size of the egg mass. The gregarious parasitoids have many eggs that take up more space than a single egg from the solitary parasitoids. This might make gregarious groups of cocoons easier to find.
To test this, the scientists first looked at how often hyperparasitoids found cocoons from each species. If a hyperparasitoid laid eggs in a cocoon (either a solitary one or one of a gregarious clutch), the cocoon was scored as a 1. If the cocoon wasn't hyperparasitized, the cocoon was scored as a 0. Again, this creates a binomial data set (either 1 or 0). To handle this, they used the Generalized Linear Model again.
Second, we tested whether hyperparasitoids exert different levels of parasitism of gregarious and solitary primary parasitoids. Within each of the years, for each Cotesia species we counted the total number of emerging wasps for each of the primary parasitoid and hyperparasitoid species. The data on species composition and their abundance per Cotesia species were subjected to Chi-square tests to assess parasitoid community differences for primary parasitoid brood size (solitary or gregarious). One of the hyperparasitoid species (i.e.,Baryscapus galactopus) parasitizes the larvae of Cotesia when these are still inside a caterpillar and lay several eggs within a single Cotesia larva. The B. galactopus brood develops when the Cotesia larvae spin their cocoon outside the caterpillar and B. galactopus wasps emerge from the Cotesia pupa. The brood size of B. galactopus on Cotesia is on average eight B. galactopus per Cotesia pupa, and therefore, we recalculated the total incidence of parasitization of Cotesia pupae by dividing the B. galactopus numbers by 8 and rounding off to the nearest whole number (numbers of each parasitoid species are presented in Table 5). All statistical tests were performed with the statistical software package Gen Stat (10th edition).
Then they tested if hyperparasitoids parasitized one species of parasitoid more than the other. During each year of the study, they counted how many parasitoid cocoons were parasitized.
But wait. The hyperparasitoids lay eggs inside the cocoon and then take it over from inside. So how could you tell which cocoons have been parasitized? By being very patient.
If scientists waited for an insect to emerge from a cocoon, they could tell if it had been parasitized or not. Image by the US Department of Agriculture.
The scientists collected all of these different cocoons or groups of cocoons (called broods above) in tiny capsules. And then they waited. Once each cocoon was done developing, a parasitoid or a hyperparasitoid would come out, or emerge. If a hyperparasitoid emerged, the parasitoid cocoon had been taken over and eaten by hyperparasitoid young.
They recorded the number and the species of hyperparasitoids that emerged from the cocoons. This data was then used in a Chi-square test.
Chi-square tests measure how what you saw fits with what you expected to see. If hyperparasitoids had no preference for a specific parasitoid species, what would the scientists expect to see?
A chi-square test compares what the scientists expected to see with what they really observed. Click for more detail.
The number of hyperparasitoid young emerging from each parasitoid cocoon type should be similar. The scientists could then compare this idea to what they really saw. If what they saw differed from this, then the wasps preferred one species.
Here, the scientists also explain a change or correction that they did to the data. There is one species of hyperparasitoid that lays an average of eight eggs in one cocoon. Many more of this species would emerge in total than other species because of this fact.
Scientists count parasitized cocoons by the number of wasps that emerged. Because of this, the species that lays eight eggs in one cocoon would increase the counts the scientists made above the real measure they wanted. To fix this, the number of young of this species that emerged was divided by eight. The new number better represented how many hyperparasitoids were choosing each cocoon type.
We used Partial Least Squares Projection to Latent Structures-Discriminant Analysis (PLS-DA) to analyze which of the compounds contributed most to describing the difference among plant treatments. The compounds that scored >1 in their Variable Importance in the Projection (VIP) scores were subjected to Mann–Whitney U tests among treatment pairs to test for significant differences among treatments.
Now back over to the plant chemicals one last time. To do the final analysis of the plant chemical data, the scientists used another type of test. They knew the chemicals the plants released differed with plant treatment. But they still wanted to see which chemicals caused them to differ.
The scientists compared chemicals from the four different plant treatments to see exactly how the volatiles differed. Image by Rajeevvadakkedath.
For this, they used a test called Partial Least Squares, though it is also called Projection to Latent Structures (PLS). Like the Chi-square test, the PLS also compares what was predicted with what was observed. But the PLS can handle more factors. Here, the scientists used it to test chemical composition against each plant treatment.
Certain chemicals were more unique than others in different treatments. This earned the chemical a higher score in the Projection test. Chemicals that were most different were then tested further. For this, the scientists used a Mann-Whitney U test, which compares two groups that have abnormal data.
Scientists can't do all of this great research alone. It takes all kinds of help from other people.
The Acknowledgments section is where they thank the people who were involved but didn't end up being authors of the article.
We thank Kees Zwakhals for identification of Mesochorus gemellus, the Center for Genetic Resources (CGN) and Bejo Zaden for providing seeds of the cultivars, Unifarm for maintenance of the plants and the field site, and Tibor Bukovinszky for the pictures of parasitoids and hyperparasitoids.
Author Contributions
Just knowing who was an author on the paper doesn't tell us which author did what during the experiment. That is what the Author Contributions section is for.
The author(s) have made the following declarations about their contributions: Conceived and designed the experiments: EHP FZ. Performed the experiments: EHP MB FZ BTW AEB. Analyzed the data: EHP BTW YJ. Wrote the paper: EHP MD JAH LEMV JJAL.
References
For most of the information included in an article, the authors need to present the citation (or reference) that shows the information is valid. Image by Webcomic xkcd.
Throughout the paper, you may have noticed numbers at the end of some sentences. If the author stated information that isn't widely known (or sometimes even if it is), they are supposed to tell you where they got that information.
Each of the numbers you saw matches with a specific article listed below. This is called a citation, and is the same thing you do when you write a Bibliography. This lets you track down where the authors found the information that they told you.
If you are interested in any of the things the author cited, you should try to look up the matching article and give it a read.
Vet LEM, Dicke M (1992) Ecology of infochemical use by natural enemies in a tritrophic context. Annu Rev Entomol 37: 141–172. doi: 10.1146/annurev.en.37.010192.001041
Hare JD (2011) Ecological role of volatiles produced by plants in response to damage by herbivorous insects. Annu Rev Entomol 56: 161–180. doi: 10.1146/annurev-ento-120709-144753
Kessler A, Heil M (2011) The multiple faces of indirect defences and their agents of natural selection. Funct Ecol 25: 348–357. doi: 10.1111/j.1365-2435.2010.01818.x
4. Price PW, Bouton CE, Gross P, Mcpheron BA, Thompson JN, Weis AE (1980) Interactions among three trophic levels: influence of plants on interactions between insect herbivores and natural enemies. Annu Rev Ecol Syst 11: 41–65. doi: 10.1146/annurev.es.11.110180.000353
5. Dicke M, van Loon JJA, Soler R (2009) Chemical complexity of volatiles from plants induced by multiple attack. Nat Chem Biol 5: 317–324. doi: 10.1038/nchembio.169
6. Dicke M, Baldwin IT (2010) The evolutionary context for herbivore-induced plant volatiles: beyond the “cry for help.”. Trends Plant Sci 15: 167–175. doi: 10.1016/j.tplants.2009.12.002
7. Heil M, Karban R (2010) Explaining evolution of plant communication by airborne signals. Trends Ecol Evol 25: 137–144. doi: 10.1016/j.tree.2009.09.010
8. Halitschke R, Stenberg JA, Kessler D, Kessler A, Baldwin IT (2008) Shared signals—“alarm calls” from plants increase apparency to herbivores and their natural enemies in nature. Ecol Lett 11: 24–34. doi: 10.1111/j.1461-0248.2007.01123.x
9. Ohgushi T (2008) Herbivore-induced indirect interaction webs on terrestrial plants: the importance of non-trophic, indirect, and facilitative interactions. Entomol Exp Appl 128: 217–229. doi: 10.1111/j.1570-7458.2008.00705.x
10. Van Loon JJA, de Boer JG, Dicke M (2000) Parasitoid-plant mutualism: parasitoid attack of herbivore increases plant reproduction. Entomol Exp Appl 97: 219–227. doi: 10.1046/j.1570-7458.2000.00733.x
11. Fritzsche-Hoballah MEF, Turlings TCJ (2001) Experimental evidence that plants under caterpillar attack may benefit from attracting parasitoids. Evol Ecol Res 3: 553–565.
12. Smallegange RC, van Loon JJA, Blatt SE, Harvey JA, Dicke M (2008) Parasitoid load affects plant fitness in a tritrophic system. Entomol Exp Appl 128: 172–183. doi: 10.1111/j.1570-7458.2008.00693.x
13. Harvey JA, van Dam NM, Gols R (2003) Interactions over four trophic levels: foodplant quality affects development of a hyperparasitoid as mediated through a herbivore and its primary parasitoid. J Anim Ecol 72: 520–531. doi: 10.1046/j.1365-2656.2003.00722.x
14. Bukovinszky T, van Veen FJF, Jongema Y, Dicke M (2008) Direct and indirect consequences of resource quality on food web structure. Science 319: 804–807. doi: 10.1126/science.1148310
15. Sullivan DJ, Völkl W (1999) Hyperparasitism: multitrophic ecology and behaviour. Annu Rev Entomol 44: 291–315. doi: 10.1146/annurev.ento.44.1.291
16. Völkl W, Sullivan DJ (2000) Foraging behaviour, host plant and host location in the aphid hyperparasitoid Euneura augarus. Entomol Exp Appl 97: 47–56. doi: 10.1046/j.1570-7458.2000.00715.x
17. Harvey JA, Wagenaar R, Bezemer TM (2009) Interactions to the fifth trophic level: secondary and tertiary parasitoid wasps show extraordinary efficiency in utilizing host resources. J Anim Ecol 78: 686–692. doi: 10.1111/j.1365-2656.2008.01516.x
18. Fatouros NE, Van Loon JJA, Hordijk KA, Smid HM, Dicke M (2005) Herbivore-induced plant volatiles mediate in-flight host discrimination by parasitoids. J Chem Ecol 31: 2033–2047. doi: 10.1007/s10886-005-6076-5
19. Poelman EH, Gols R, Snoeren TAL, Muru D, Smid HM, Dicke M (2011) Indirect plant-mediated interactions among parasitoid larvae. Ecol Lett 14: 670–676. doi: 10.1111/j.1461-0248.2011.01629.x
20. Poelman EH, Zheng S-J, Zhang Z, Heemskerk NM, Cortesero A-M, Dicke M (2011) Parasitoid-specific induction of plant responses to parasitized herbivores affects colonization by subsequent herbivores. Proc Natl Acad Sci USA 108: 19647–19652. doi: 10.1073/pnas.1110748108
21. McDonald RC, Kok LT (1991) Hyperparasites attacking Cotesia glomerata (L.) and Cotesia rubecula (Marshall) (Hymenoptera: Braconidae) in Southwestern Virginia. Biol Control 1: 170–175. doi: 10.1016/1049-9644(91)90116-h
22. Brodeur J, Geervliet JBF, Vet LEM (1998) Effects of Pieris host species on life history parameters in a solitary specialist and gregarious generalist parasitoid (Cotesiaspecies). Entomol Exp Appl 86: 145–152. doi: 10.1046/j.1570-7458.1998.00275.x
23. Le Masurier AD (1987) A comparative study of the relationship between host size and brood size in Apanteles spp. (Hymenoptera: Braconidae). Ecol Entomol 12: 383–393. doi: 10.1111/j.1365-2311.1987.tb01019.x
24. Mattiacci L, Dicke M, Posthumus MA (1995) β-Glucosidase: an elicitor of herbivore-induced plant odor that attracts host-searching parasitic wasps. Proc Natl Acad Sci USA 92: 2036–2040. doi: 10.1073/pnas.92.6.2036
25. D'Alessandro M, Turlings TCJ (2006) Advances and challenges in the identification of volatiles that mediate interactions among plants and arthropods. Analyst 131: 24–32. doi: 10.1039/b507589k
26. Harvey JA (2000) Dynamic effects of parasitism by an endoparasitoid wasp on the development of two host species: implications for host quality and parasitoid fitness. Ecol Entomol 25: 267–278. doi: 10.1046/j.1365-2311.2000.00265.x
27. Adamo SA, (1997) How parasites alter the behavior of their insect hosts. In: Parasitic effects on host hormones and behavior, Beckage NE (Ed.). New York: Chapman & Hall. pp. 231–245.
28. Godfray HCJ (1994) Parasitoids: behaviour and evolutionary ecology. Princeton, NJ: Princeton University Press.
29. Harvey JA, Jervis MA, Gols R, Jiang N, Vet LEM (1999) Development of the parasitoid,Cotesia rubecula (Hymenoptera: Braconidae) in Pieris rapae and Pieris brassicae(Lepidoptera: Pieridae): evidence for host regulation. J Insect Physiol 45: 173–182. doi: 10.1016/s0022-1910(98)00113-9
30. Harvey JA (2008) Comparing and contrasting development and reproductive strategies in the pupal hyperparasitoids Lysibia nana and Gelis agilis (Hymenoptera: Ichneumonidae). Evol Ecol 22: 153–166. doi: 10.1007/s10682-007-9164-x
31. Harvey JA, Wagenaar R, Gols R (2011) Differing host exploitation efficiencies in two hyperparasitoids: when is a “match made in heaven”? J Insect Behav 24: 282–292. doi: 10.1007/s10905-010-9254-4
32. Poelman EH, Oduor AMO, Broekgaarden C, Hordijk CA, Jansen JJ, et al. (2009) Field parasitism of caterpillars on Brassica oleracea plants are reliably predicted by differential attraction of Cotesia parasitoids. Funct Ecol 23: 951–962. doi: 10.1111/j.1365-2435.2009.01570.x
33. Poelman EH, van Loon JJA, Dicke M (2008) Consequences of variation in plant defense for biodiversity at higher trophic levels. Trends Plant Sci 13: 534–541. doi: 10.1016/j.tplants.2008.08.003
34. Meyer KM, Vos M, Mooij WM, Gera Hol WH, Termorshuizen AJ, et al. (2009) Quantifying the impact of above- and belowground higher trophic levels on plant and herbivore performance by modeling. Oikos 118: 981–990. doi: 10.1111/j.1600-0706.2009.17220.x
35. Orre GUS, Wratten SD, Jonsson M, Hale RJ (2010) Effects of an herbivore-induced plant volatile on arthropods from three trophic levels in brassicas. Biol Control 53: 62–67. doi: 10.1016/j.biocontrol.2009.10.010
36. Geervliet JBF, Verdel MSW, Snellen H, Schaub J, Dicke M, et al. (2000) Coexistence and niche segregation by field populations of the parasitoids Cotesia glomerata and C. rubecula in the Netherlands: predicting field performance from laboratory data. Oecologia 124: 55–63. doi: 10.1007/s004420050024
37. Takabayashi J, Dicke M (1992) Response of predatory mites with differing rearing histories to volatiles of uninfested plants. Entomol Exp Appl 64: 187–193. doi: 10.1111/j.1570-7458.1992.tb01608.x
By volunteering, or simply sending us feedback on the site. Scientists, teachers, writers, illustrators, and translators are all important to the program. If you are interested in helping with the website we have a Volunteers page to get the process started.





 The listed authors of an article are the people who did most of the work. Usually, the first author (here, Erik H. Poelman) was the most involved. The first author is the person you should contact if you have questions about the article. Sometimes, the last author is the senior researcher associated with the work (here, Marcel Dicke).
The listed authors of an article are the people who did most of the work. Usually, the first author (here, Erik H. Poelman) was the most involved. The first author is the person you should contact if you have questions about the article. Sometimes, the last author is the senior researcher associated with the work (here, Marcel Dicke). Some scientific articles have an Author Summary section. If they do, it’s best to read that section first (even if it is after the Abstract).
Some scientific articles have an Author Summary section. If they do, it’s best to read that section first (even if it is after the Abstract).











 Before going to the next full section, it’s good to briefly look over some of the figures. This helps you get a better picture in your head of the information included in the paper and of what the scientists did. When you first review the images, it's good to focus on just one or two that help set up the ideas behind the experiment.
Before going to the next full section, it’s good to briefly look over some of the figures. This helps you get a better picture in your head of the information included in the paper and of what the scientists did. When you first review the images, it's good to focus on just one or two that help set up the ideas behind the experiment.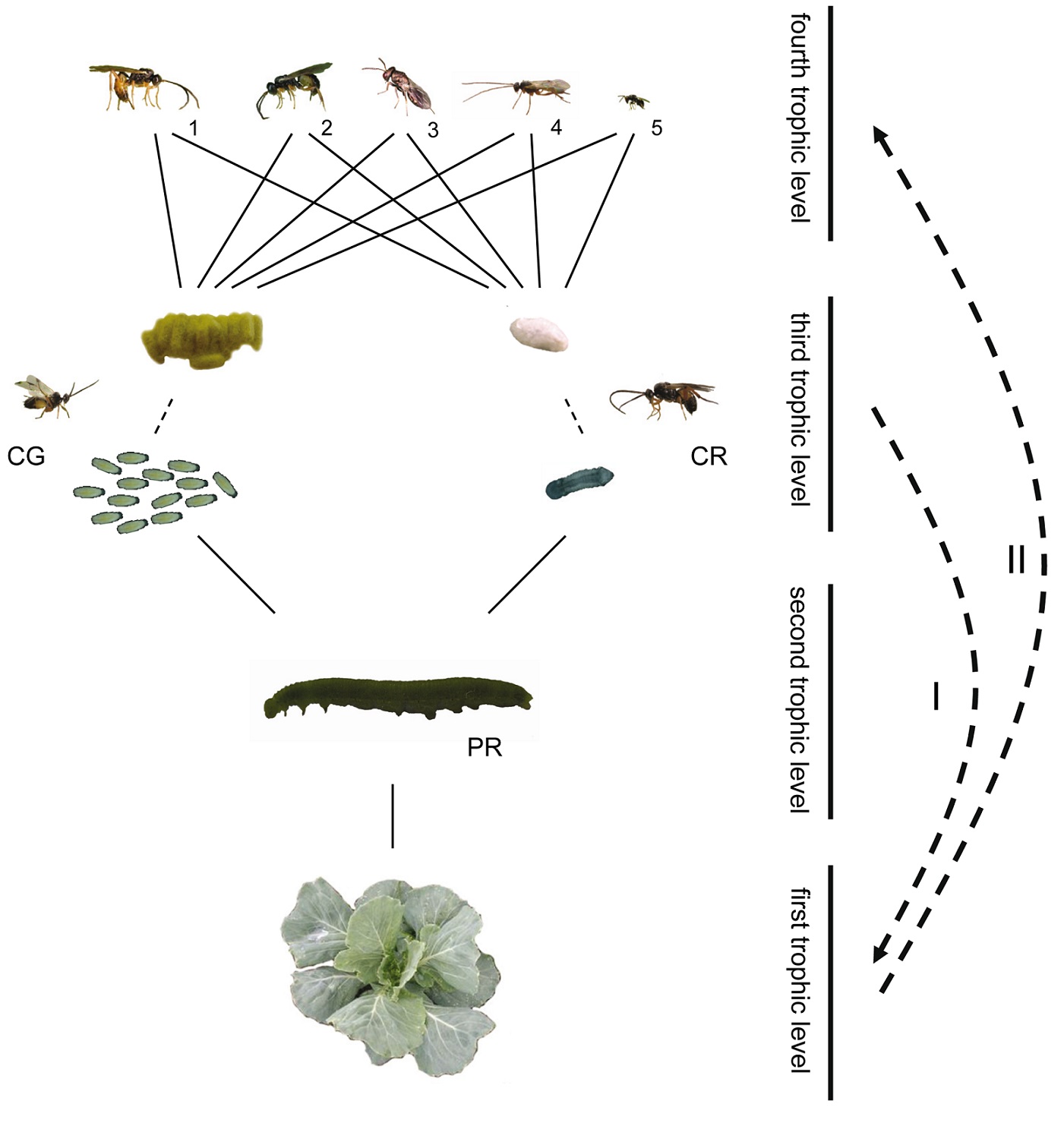

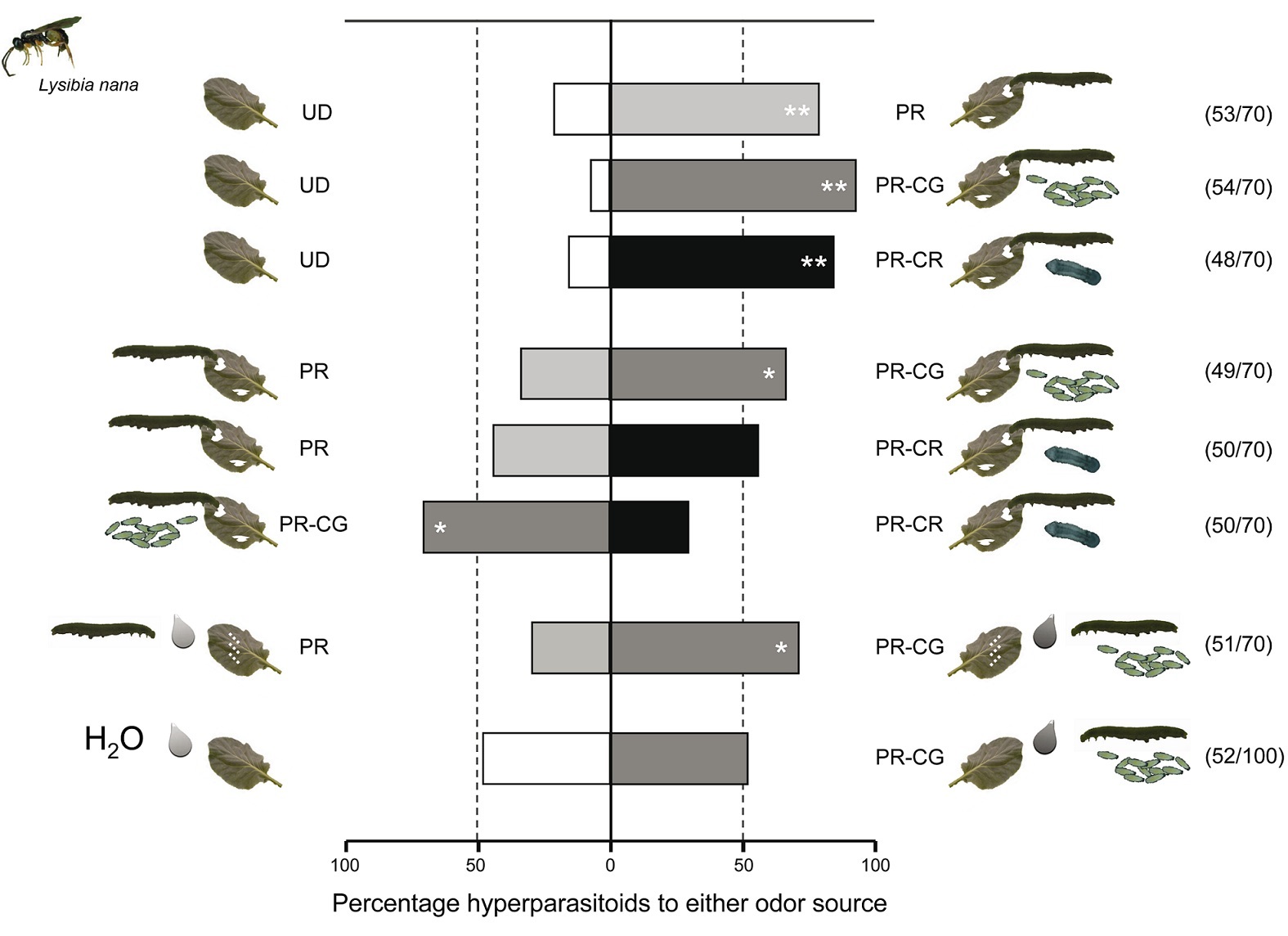




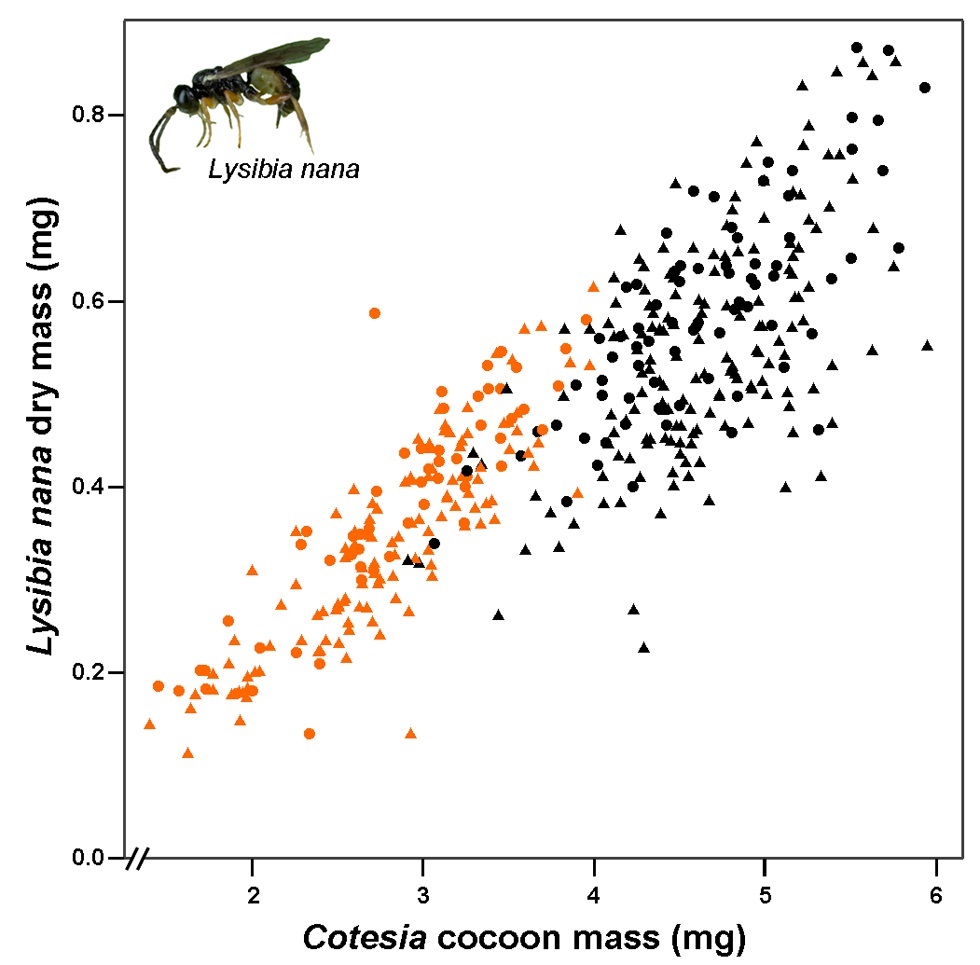

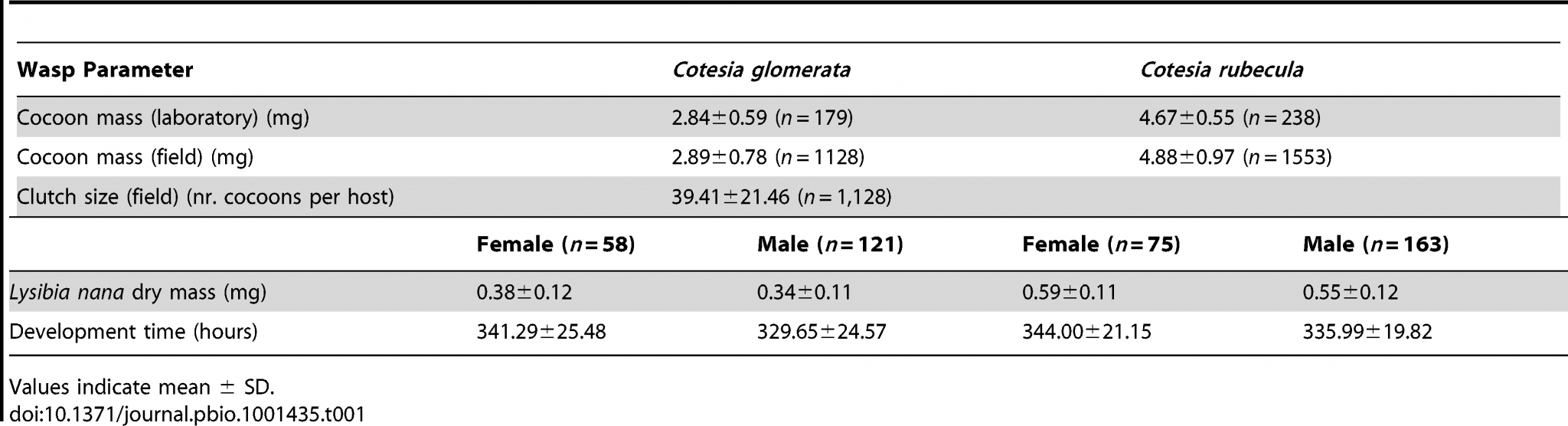

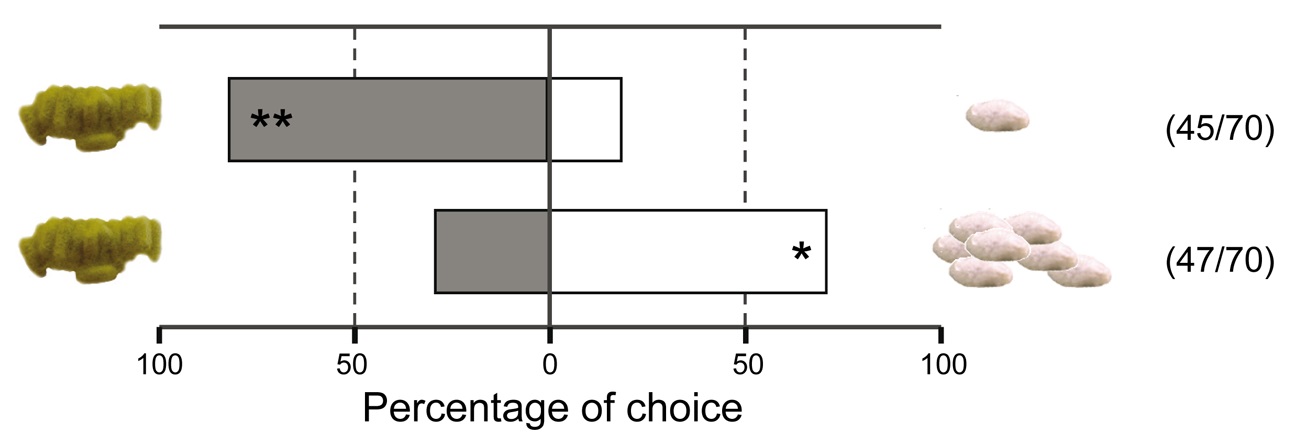








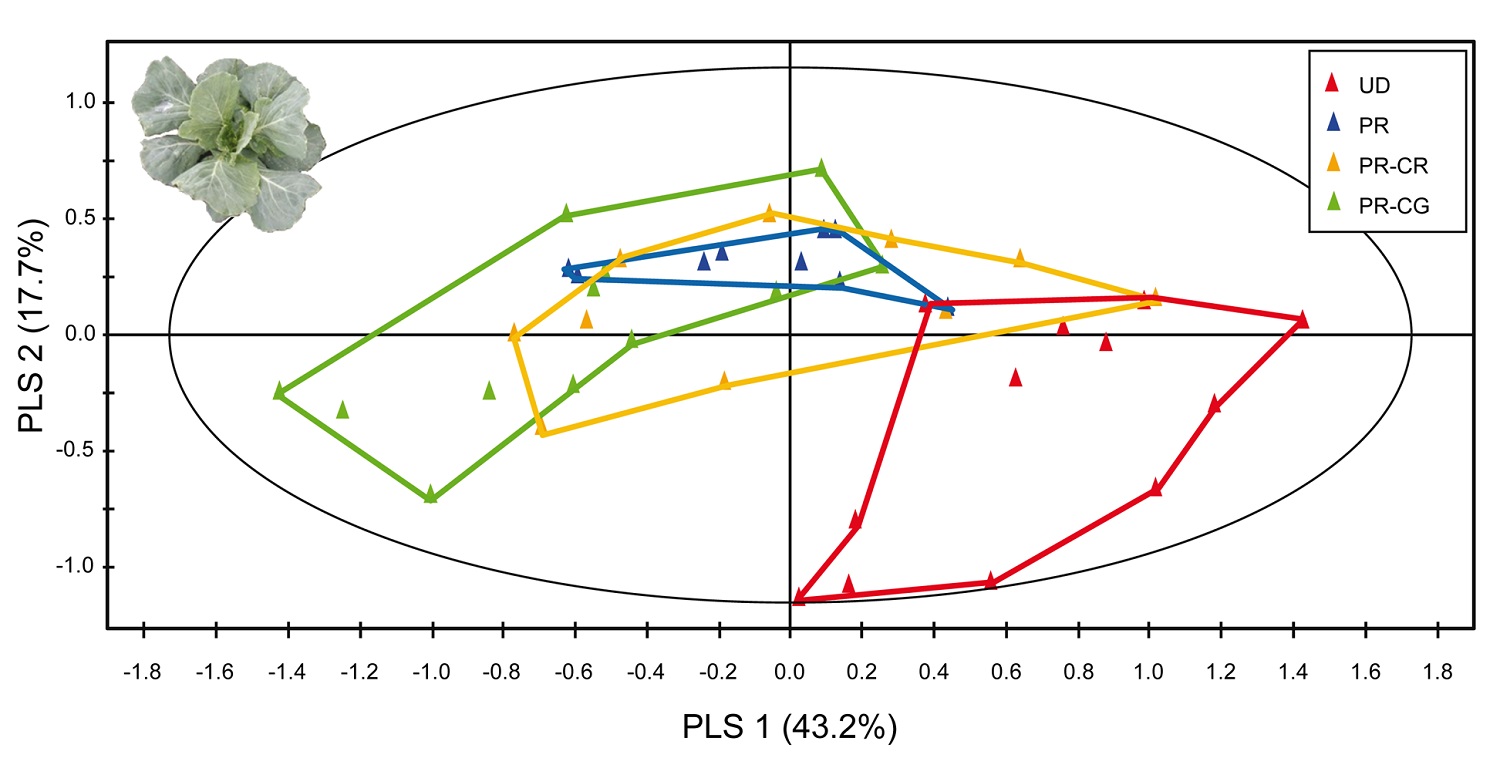

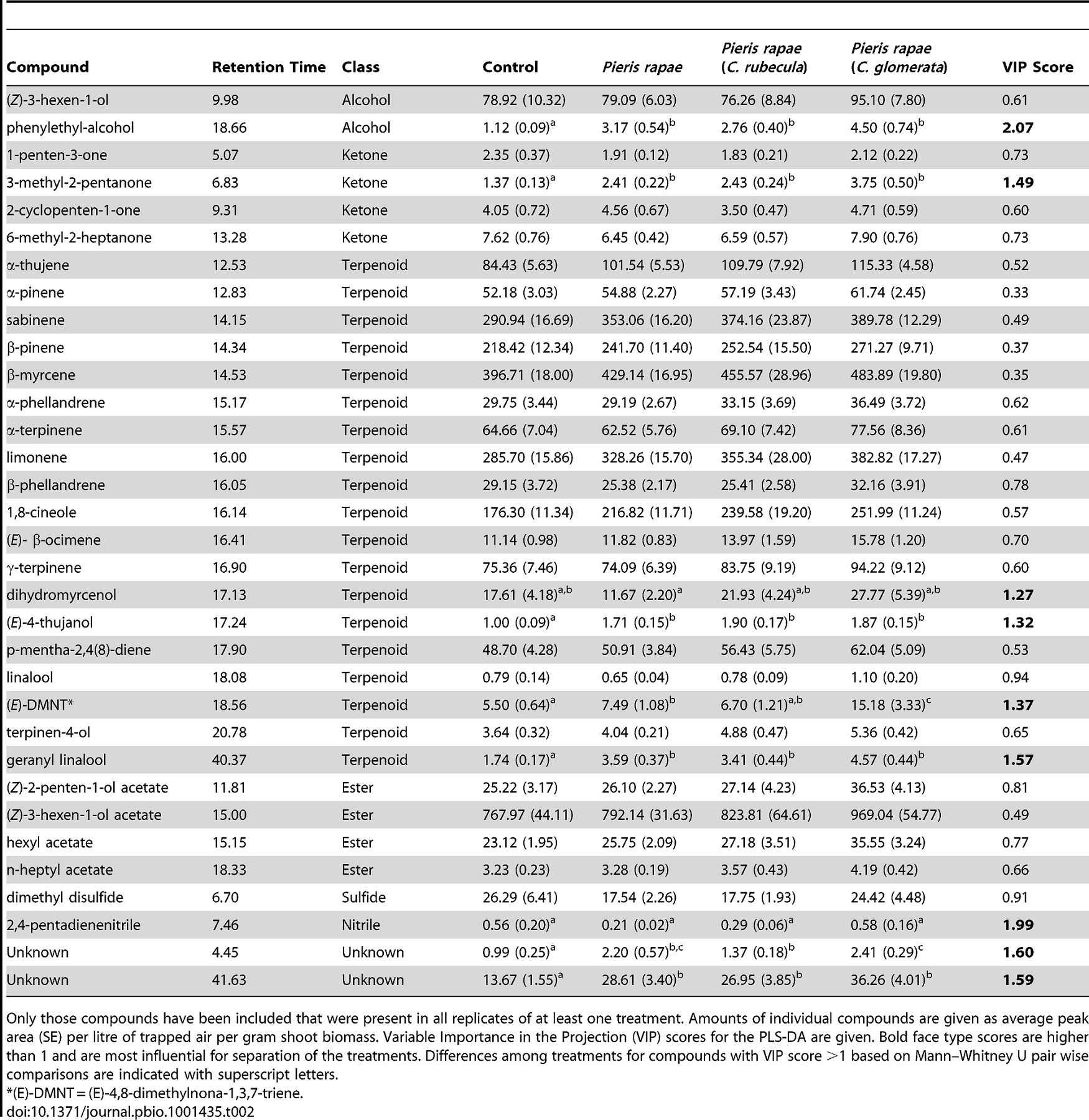



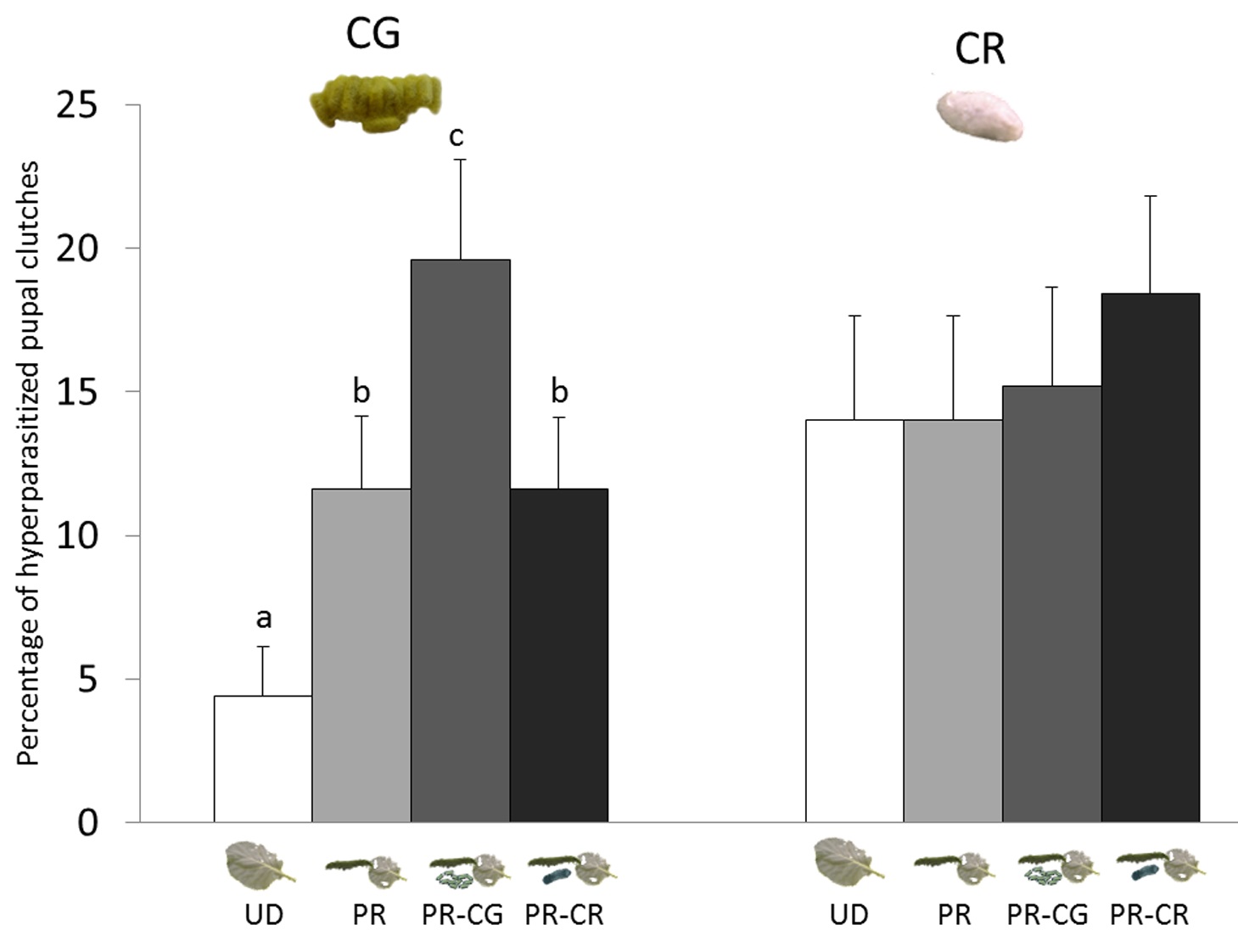
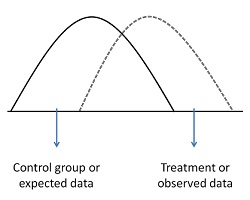


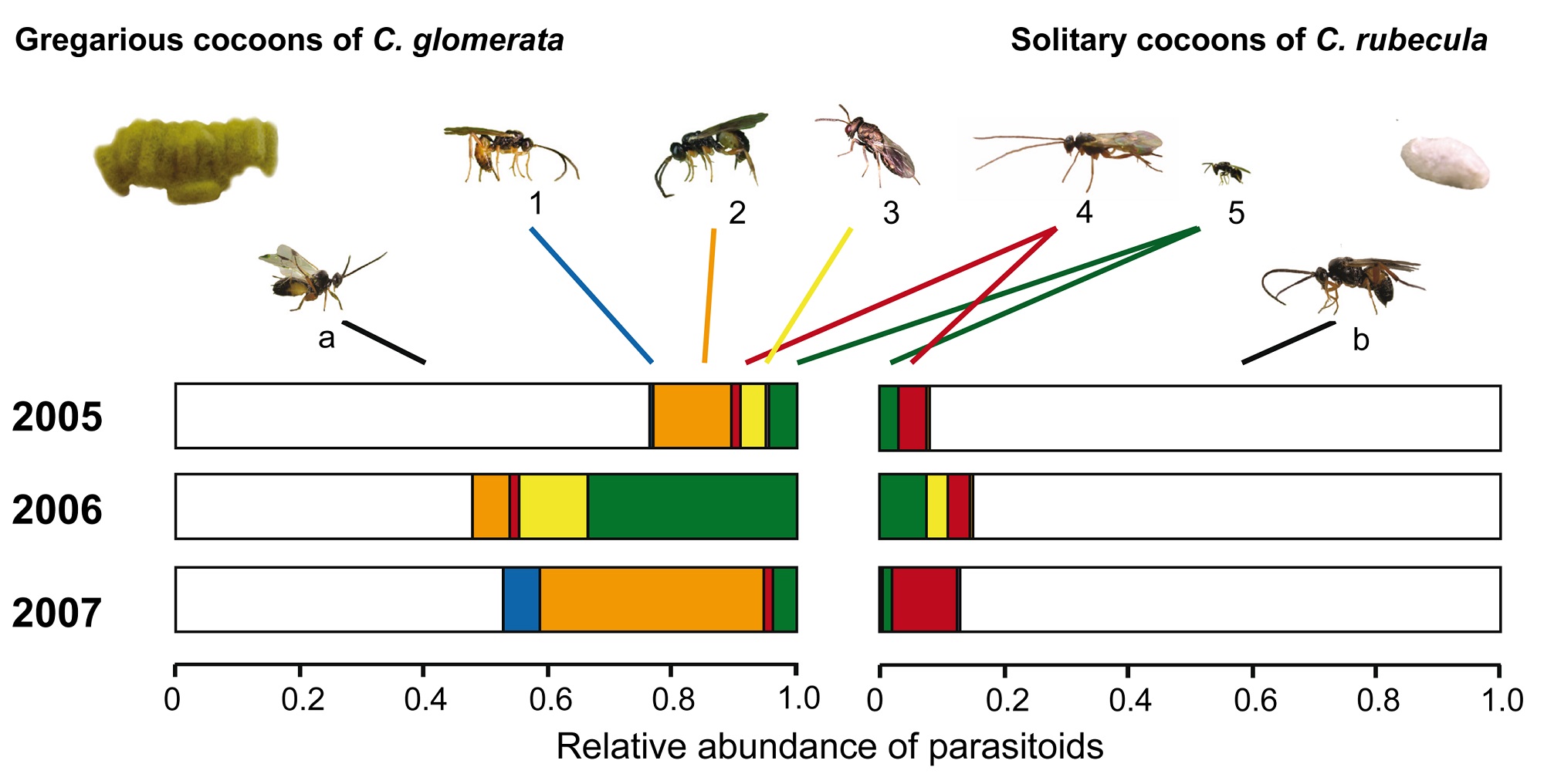

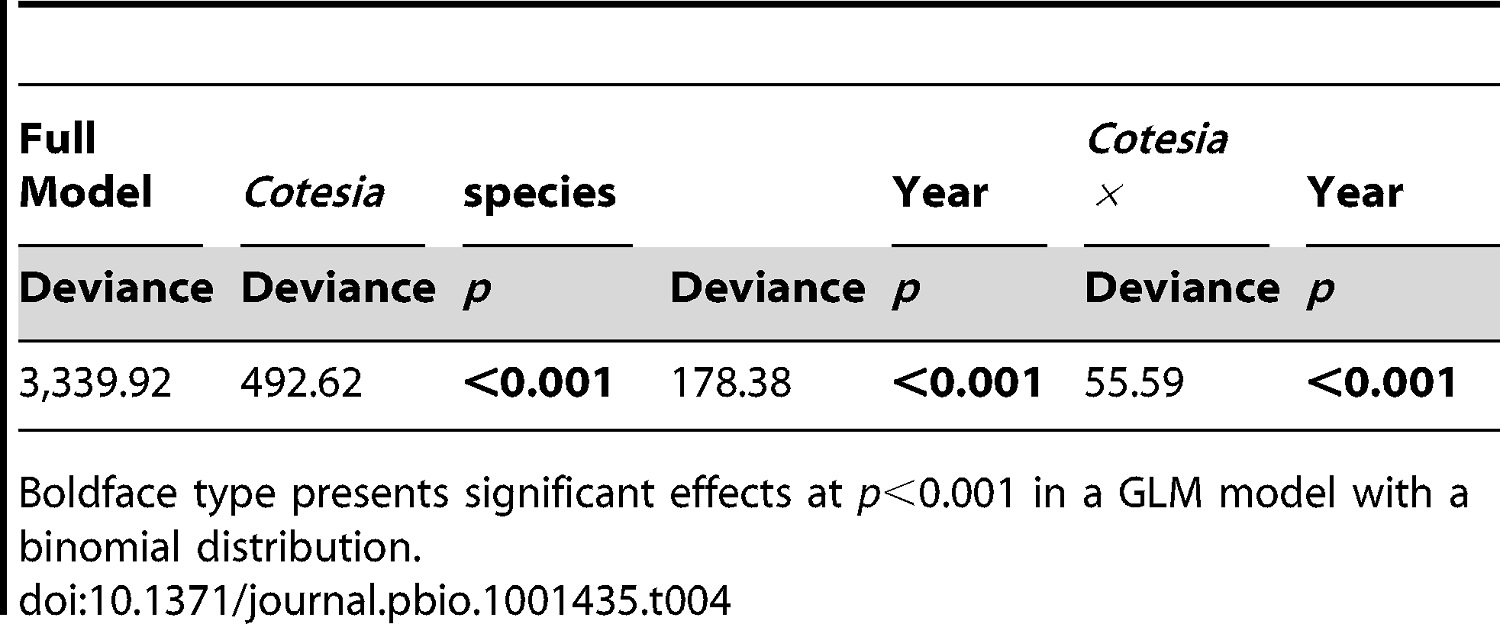

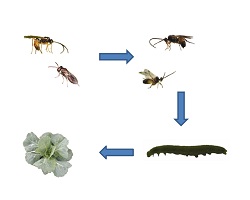








 Sometimes we save the best for last. If an article has a conclusion, this is where the authors summarize all the important findings from the whole study. This is the best section to read if you want a shorter description of the overall importance of the study.
Sometimes we save the best for last. If an article has a conclusion, this is where the authors summarize all the important findings from the whole study. This is the best section to read if you want a shorter description of the overall importance of the study.